免疫系
免疫系(めんえきけい、英語: immune system)とは、生体内で病原体などの非自己物質やがん細胞などの異常な細胞を認識して殺滅することにより、生体を病気から保護する多数の機構が集積した機構である。精密かつダイナミックな情報伝達を用いて、細胞、組織、器官が複雑に連係している。この機構はウイルスから寄生虫まで広い範囲の病原体を感知し、作用が正しく行われるために、生体自身の健常細胞や組織と区別しなければならない。
この困難な課題を克服して生き延びるために、病原体を認識して中和する機構が一つならず進化した。細菌のような簡単な単細胞生物でもウイルス感染を防御する酵素系をもっている。その他の基本的な免疫機構は古代の真核生物において進化し、植物、魚類、ハ虫類、昆虫に残存している。これらの機構はディフェンシンと呼ばれる抗微生物ペプチドが関与する機構であり、貪食機構であり、[1]補体系である。ヒトのような脊椎動物はもっと複雑な防御機構を進化させた。脊椎動物の免疫系は多数のタイプのタンパク質、細胞、器官、組織からなり、それらは互いに入り組んだダイナミックなネットワークで相互作用している。このようないっそう複雑な免疫応答の中で、ヒトの免疫系は特定の病原体に対してより効果的に認識できるよう長い間に適応してきた。この適応プロセスは適応免疫あるいは獲得免疫(あるいは後天性免疫)と呼ばれ、免疫記憶を作り出す。特定の病原体への初回応答から作られた免疫記憶は、同じ特定の病原体への2回目の遭遇に対し増強された応答をもたらす。獲得免疫のこのプロセスがワクチン接種の基礎である。
免疫系が異常を起こすと病気になる場合がある。免疫系の活動性が正常より低いと、免疫不全病が起こり感染の繰り返しや生命を脅かす感染が起こされる。免疫不全病は、重症複合免疫不全症のような遺伝病の結果であったり、レトロウイルスの感染によって起こされる後天性免疫不全症候群 (AIDS) や医薬品が原因であったりする。反対に自己免疫病は、正常組織に対しあたかも外来生物に対するように攻撃を加える、免疫系の活性亢進からもたらされる。ありふれた自己免疫病として、関節リウマチ、I型糖尿病、紅斑性狼瘡がある。免疫学は免疫系のあらゆる領域の研究をカバーし、ヒトの健康や病気に深く関係している。この分野での研究をさらに推し進めることは健康増進および病気の治療にも期待できる。
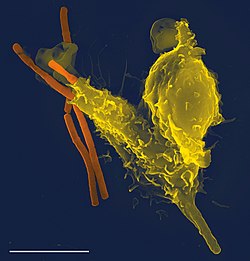
1個の好中球(黄色)が炭疽菌(オレンジ)を呑み込んでいる走査電子顕微鏡写真

走査型電子顕微鏡(SEM)による画像。Tリンパ球(右)、血小板(中央)、赤血球(左)フレデリック国立癌研究所
目次
1 概要
1.1 重層的防御
2 表面(上皮)の防壁
2.1 機械的防壁
2.2 化学的防壁
2.3 生物学的防壁
3 自然免疫
3.1 体液性すなわち化学的防壁
3.1.1 炎症
3.1.2 補体系
3.2 細胞による防壁
4 特異的・適応的な獲得免疫
4.1 リンパ球
4.1.1 細胞傷害性T細胞(CTL)
4.1.2 ヘルパーT細胞 (Th)
4.1.3 γδT細胞
4.1.4 B細胞と抗体
4.1.5 代替的適応免疫系
4.2 免疫記憶
4.2.1 受動的な記憶
4.2.2 能動的な記憶と免疫処置
5 ヒトの免疫異常
5.1 免疫不全
5.2 自己免疫
5.3 過敏症
6 他の機構
7 腫瘍免疫
8 生理学的制御
9 医学における操作
10 病原体の操作
11 免疫学とその歴史
12 脚注・出典
13 関連項目
14 外部リンク
概要
免疫とは、ヒトや動物などが持つ、体内に入り込んだ「自分とは異なる異物」(非自己)を排除する、生体の恒常性維持機構の一つである。一般に、薬物や化学物質などの排除には、肝臓の酵素による代謝が働くのに対し、免疫はそれよりも高分子であるタンパク質(ヘビ毒やハチ毒など)や、体内に侵入した病原体を排除するための機構として働くことが多い。特に病原体による感染から身を守るための感染防御機構として重要であり、単に「免疫」と呼ぶ場合には、この感染防御免疫のことを指す場合も多い。
免疫系は自然免疫(先天性免疫、基本免疫)と獲得免疫(後天性免疫、適応免疫)とに大別される。
自然免疫にはある特殊な細胞が備わっており、それらは侵入物が自己を再生産したり宿主に対し重大な被害をもたらす前に発見、排除し、病原体が体内で増殖して宿主に深刻な害を及ぼす前に対処する事ができる。
一方の獲得免疫は抗体や補体などの血中タンパク質による体液性免疫の他に、リンパ球などの細胞による細胞性免疫によって担われている。リンパ球には分化成熟して免疫グロブリンを産生するB細胞のほかに、胸腺で分化成熟するT細胞などがある。その他、食作用によって抗原を取り込んで分解してT細胞に提示する樹状細胞なども免疫機能の発現に関与する。これらの細胞は骨髄で産生され、胸腺やリンパ節、脾臓などのリンパ系組織での相互作用をへて有効な機能を発揮するようになる。
自然免疫も適応免疫もその効果のほどは自己と非自己の分子の区別ができる能力をもった免疫系かどうかにかかっている。
免疫学において自己分子とは、免疫系によって外来物質と区別できるような自己の身体要素のことである[2]。
反対に非自己分子とは、免疫系によって外来物質と区別される外来分子のことである。非自己分子の一つのクラスは、抗原(antigen; これはantibody generatorの短縮語である)と呼ばれ、特異的な免疫受容体に結合し、免疫応答を誘発する物質と定義される[3]。
| 項目 | 自然免疫 | 獲得免疫 |
|---|---|---|
病原体と抗体の反応 | 非特異的 | 特異的 |
| 接触後最大応答までの時間 | 短い(即座) | 長い(病原体に適応するための遅延) |
| 関与する成分 | 細胞性および体液性 | 細胞性および体液性 |
| 細胞の名称 | 白血球 | リンパ球 |
| 免疫記憶 | なし | あり |
| 生物界での分布 | ほとんど全ての生物 | 顎をもった脊椎動物 |
重層的防御
免疫系は、感染から生体を、特異性を高めながら重層的な防御体制で守る。最も簡単なのは、物理的な障壁で、細菌やウイルスが生体に侵入するのを防ぐことである。
病原体がこの障壁を突破して体内に侵入したとき、即座に自然免疫(先天性免疫とも呼ばれる)はそれを感知し非特異的に対応して排除する。
自然免疫はあらゆる植物および動物に認められる[4]。
しかし病原体が自然免疫もうまく逃れたなら脊椎動物は第3階層の防御反応を繰り出す。これが獲得免疫であり、一度感染源に接触することで自然免疫によって発動される。
後天性免疫系は病原体を認識して攻撃するが、感染を受ける間、応答を病原体への認識が改善されるよう適応する。
この機構は、病原体が排除された後も免疫記憶として残り、次いで、同一(あるいは非常に似通った)の病原体に遭遇する度に強化される仕組みになっていて、より早く強力な攻撃が加えられる[5]。
表面(上皮)の防壁
病原体に曝露された細胞上皮(いわゆる皮膚だけでなく、粘膜や腸管などを含む)には生体を感染から守る障壁があり、機械的、化学的、生物学的に守っている。
機械的防壁
葉に見られるワックス性クチクラ、昆虫の外骨格、産み落とされた卵の殻や膜、そして皮膚。これらは機械的障壁の例であって、感染に対する防御ラインの第一線にある[3]。皮膚は上皮、外層、真皮から構成され、ほとんどの感染因子を物理的に遮断する。
しかし生体は外界に対して完全に密閉されることはできないのだから、肺、腸、性尿器路など、外界に対して開口された部分を守るのには他の系を作動させる。肺と気管では、咳やくしゃみの気流と繊毛の動きによって、病原体や呼吸管の他の刺激物を機械的に排除する。涙や尿を流す行動も病原体を機械的に追い出すし、呼吸器の呼吸管や消化器の唾液や胃腸管から分泌される粘液には、微生物を捕らえ絡み取る働きがある[6]。
化学的防壁
化学的障壁も感染防御に働く。皮膚は、ケラチンを豊富に含む細胞がきっちり密に並んで構成されている。これが水を弾き、皮膚を弱酸性に保つため、皮膚はバクテリアの増殖を抑える化学的障壁としても働く。
皮膚や呼吸管はβ-ディフェンシンのような抗微生物ペプチドを分泌する[7]。
唾液、涙、母乳に含まれるリゾチームやホスホリパーゼA2等の酵素も抗菌作用がある抗細菌物質である[8][9]。
膣分泌液は初経後のわずかにでも酸性に傾いたとき化学的障壁として働くし、精液は病原体殺滅性のあるスペルミンやディフェンシンや亜鉛を含む[10][11]。
分泌液として胃液には胃酸が極端な低pHを示すとともに消化酵素のタンパク質分解酵素を含んでおり、摂取された病原体に対して強力な化学的防御の働きがある。
生物学的防壁
生殖尿管や胃腸管では共生している細菌叢が病原菌と養分や繁殖場所をめぐって病原体と競争して生物学的障壁として機能する。この場合、pHや利用できるイオンのような環境を変えることもある[12]。このことは病原体が発症可能な個体数まで増殖できる可能性を減らす。腸内では、腸内細菌が働いている。ヨーグルトに通常含まれている乳酸菌のような純粋培養によって良性細菌叢を再導入することは、子供たちの腸管感染での微生物集団のバランスを健康なものに保つのを助ける働きがあるという証拠が出されている。これは細菌性胃腸炎、炎症性腸疾患、尿路感染症、術後感染の研究の予備的データに希望を与えている[13][14][15]。
細菌性の感染症に対してはしばしば抗生物質が用いられるが、大部分の抗生物質は病原体となる細菌と正常な細菌の両方に非特異的に作用するし、カビには効かないので、抗生物質の経口投与によって、真菌を異常に増殖させ、膣カンジダ症のような真菌症を引き起こす場合がある[16]。
自然免疫
病原体が上皮での防壁を突破し、微生物や毒性物質が生体内にうまく侵入できると、続いて生体内の自然免疫系(先天性免疫系)と対峙する。これは体液的・化学的・細胞的な障壁による宿主の保護機構である。白血球やリンパ球などの細胞や機構が動員されて宿主を守り、その際に通常は炎症反応が起きる。
この自然免疫応答は普通生体が微生物を構造パターン認識受容体で感知するときに発動する。この構造パターンは広い範囲の微生物グループの間で保存されている[17]。
あるいは細胞は障害を受けると警戒シグナルを出す。それらの全てではないが多くは病原体を認識する同じ受容体によって感知される[18]。
自然免疫系による防御は非特異的であり、病原体に対して包括的な応答を行う[3]。つまり様々な病原体に対して別個に応答するのではなく常に汎用的な方法で対処するがゆえに、効力を発動するまでの時間が短く、いわば常に臨戦態勢にある。反面、獲得免疫系のような免疫記憶が無く、長期にわたって防御する仕組みではない。
自然免疫系は大部分の生物にとって宿主防御の主要な系であり[4]、植物・菌類・昆虫・多細胞生物(哺乳類などの高等脊椎動物を除く)においては主要な防御システムである。
原始的な生命も持っており、進化的に古い防御方法であると考えられている。また、Toll様受容体、Nodタンパク質、RIG-I(病原微生物に対するセンサー)などの研究が20世紀末から進展し、自然免疫が高等動物にも存在するのみならず、獲得免疫が成立する前提として重要なメカニズムである(たとえばマクロファージや樹状細胞が病原体の存在により直接活性化される)ことが明らかとなった。
体液性すなわち化学的防壁
炎症
炎症は病原体の感染や刺激に対し免疫系が最初に起こす応答の一つである[19]。
炎症の徴候は、発赤、疼痛、熱感、腫脹の4つで、組織に流入する血液の増加によって起こされる。
炎症は傷害や感染を受けた細胞が分泌するエイコサノイドとサイトカインと呼ばれる特定の化学伝達物質群などの化学的因子によって起こり、感染に対する免疫機構の正常な反応である。
エイコサノイドにはプロスタグランジンが含まれ、この物質は炎症に関係した場合、発熱と血管拡張を起こす。
また同じくエイコサノイドに含まれるロイコトリエンはある種の白血球(リンパ球)に作用する[20][21]。
サイトカインの種類には白血球間の情報伝達に関与するインターロイキン、走化性を促してマクロファージなどを呼び寄せるケモカイン、宿主細胞のタンパク質合成を停止させるようなウイルスに対して、抗ウイルス活性をもったインターフェロンなどがある[22]。
増殖因子や細胞毒性因子も分泌される場合がある。
これらのサイトカインや他の化学物質は免疫細胞を感染部位に動員し、病原体を排除してから損傷を受けたあらゆる組織の修復を促す[23]。
補体系
補体系は、外来細胞の表面に攻撃を加える、或は他の細胞によって破壊されるよう指標を付けるための抗体能力を補助(補う)する生化学的カスケード(連鎖)である。補体とは、抗体の機能を補助、あるいは補完するたんぱく質である。
20以上のタンパク質が関与し、抗体による病原体殺滅を補 強する能力をもつ、という意味で名づけられた。補体は自然免疫応答において主要な体液性要素をなす[24][25]。
補体系をもつ種は数多く、哺乳類に限らず、植物、魚類、無脊椎動物の一部など、ある程度原始的な生物でも持ち合わせている[26]。
ヒトではこの応答はこれら微生物に付着した抗体に補体が結合することにより、あるいは微生物の表面の炭水化物に補体タンパク質が結合することにより活性化される。この認識シグナルが、速やかな殺滅応答を発動する[27]。応答のスピードを決めるのは引き続いて起こる補体分子のタンパク質分解の活性化によって起こるシグナル増強の程度である。
多くの補体タンパク質はタンパク質分解的切断によって活性化されるとプロテアーゼ(タンパク質分解酵素)に成る。
補体タンパク質が微生物に付着した後、補体自身のタンパク質分解酵素活性が発現し、続いて他の補体タンパク質分解酵素が活性化され、これが連続して起こる。これは触媒反応カスケードを引き起こし、最初のシグナルを正のフィードバックでコントロールしながら増強するものである[28]。補体がこうしてまとわりつくことによって細胞膜は破壊され病原体は殺される[24]。
補体の活性化によりしばしば感染細胞の原形質破壊が起こり、それによる感染細胞の細胞溶解、病原体の死を引き起こす。
補体機構の最終産物C5b6789は別名細胞膜障害性複合体とも言い、感染した細胞や感染源の細胞膜を破壊することで、サイトリシスや溶菌を起こす。これを免疫溶菌現象、あるいは免疫溶菌反応といい、細菌への防御においては好中球の貪食と並び重要な機構である。
カスケードはペプチドを産生して免疫細胞を誘引し、血管の透過性を更新し、病原体の表面をオプソニン化(コート)して破壊できるようマークを付ける。
補体系は病原体表面をオプソニン化(或はコーティング)することで、病原体が他の細胞に破壊されるよう札(タグ,tag)を付け、炎症細胞の回復を誘発し、中和された抗原抗体複合体の残骸を除去する。
細胞による防壁

正常なヒトの循環血液の、走査型電子顕微鏡(SEM)画像。赤血球および突起物で覆われでこぼこした少数のリンパ球を含んだ白血球を認め、ほかに単球、好中球、そして多数の小さな板状の血小板を認める。
白血球はどの器官や組織とも結合しているのではなく、単一の細胞からなる器官であり、独立した単細胞生物のように行動する、自然免疫系の右腕である[3]。自然免疫系の白血球には、肥満細胞、好酸球、好塩基球、ナチュラルキラー細胞、食細胞(マクロファージ、好中球、樹状細胞)や感染誘引可能性病原体を識別する機能がある。これらの細胞は病原体を認識し排除するが、微生物を呑み込んで殺するか、より大きな病原体に対しては接触して攻撃する[26]。自然免疫は感染の最初の段階で働くが、多くの感染源は自然免疫を回避するための戦略を発達させてきた。自然免疫系細胞は更に特異的適応的な獲得免疫に於いては重要なメディエーターであり[5]、抗原提示として知られる過程を通すことでそれを活性化することが出来る。
貪食機能は細胞性自然免疫で重要な役割をもっており、病原体や粒状物を呑み込み食す食細胞と呼ばれる細胞によって行われる。食細胞は感染源や粒子を貪食、すなわち食うことによって排除する役割を担う。食細胞は普段は体内を巡回して病原体を探しているが、サイトカインによって特定の部位に誘導される[3]。病原体は一旦食細胞に呑み込まれるとファーゴソームと呼ばれる細胞内小胞によって捕らえられ、続いてリソソームと呼ばれる今一つ別の小胞と融合してファーゴリソソームを形成する。病原体は消化酵素によって、あるいは呼吸バーストに続くフリーラジカルのファーゴリソソームへの放出によって殺滅される[29][30]。貪食機能は栄養素獲得のために進化したが、食細胞ではこの役割が拡張されて病原体の貪食を含んだ防御機構として働く[31]。貪食機能は、食細胞が脊椎動物にも無脊椎動物にも存在することから、おそらく宿主防御の最も古い形を示したものであろう[32]。
好中球とマクロファージは侵入病原体を捜して体内全体を移動している食細胞である[33]。マクロファージ上や好中球上のレセプターにバクテリア分子が結合するとバクテリアの貪食や破壊が始まる。
- 好中球
- 通常血流中に存在し、食細胞の中で最も数が多い。通常全循環白血球の50%〜60%を占める[34]。特に細菌感染の結果生じる炎症急性期には好中球は走化性というプロセスによって炎症部位に移動する。大抵の場合、感染の生じた現場に最初に到着する細胞である。
マクロファージ(大食細胞)- 組織中に存在し、侵入した感染源を追って組織や細胞間スペースにも入れる。多才な細胞で、酵素、補体タンパク質、それにインターロイキン-1のような制御因子など広範囲にわたる化学物質を産生する[35]。マクロファージは死体・ゴミあさりの(スカベンジャー)細胞としても働き、体内の役に立たなくなった細胞、およびその他の崩壊沈着物の除去および適応免疫系を活性化する抗原提示細胞として働く[5]。
樹状細胞(DC; dendritic cell)- 外界に接する組織の中に存在する食細胞である。したがってこの細胞は主に皮膚、鼻、肺、胃、腸に存在する[36]。この細胞の名称は神経細胞の樹状突起(dendrite)に似ていることから付けられた。神経細胞も樹状細胞も樹状突起を多数もっているが、神経機能には関与していない。樹状細胞は適応免疫系の鍵となるT細胞に抗原を提示するので、自然免疫と獲得免疫の橋渡しをしている[36]。
ナチュラルキラー細胞(NK細胞)
腫瘍細胞やウイルス感染症腺細胞を非特異的に攻撃して破壊する[37](ちなみにこれは炎症反応には含まない)。
好塩基球と好酸球- 好中球と関係があり、寄生虫に対する防御の際に化学メディエータを分泌する。また、喘息などのアレルギーにも関与する[38]。
肥満細胞(マスト細胞)
結合組織や粘膜に存在し、感染防御や傷の回復、炎症応答を制御する[39]。この細胞は最も多くはアレルギーとアナフィラキシーに関与する[34]。
特異的・適応的な獲得免疫
獲得免疫系は初期の脊椎動物に進化し、より強力な免疫反応を起こし、個々の病原体が特定の型であることを示す抗原によって判別し記憶(免疫記憶)する機構である[40]。応答は抗原特異的であり、抗原提示と呼ばれるプロセスの間に特異的な非自己の抗原であるという認識が行われる必要がある。抗原特異性の認識によって、特定の病原体あるいは特定の病原体感染細胞に対して調整された応答の発動を可能とする。このような調整された応答を開始する能力は体内の記憶細胞によって保持される。もし病原体が1回以上生体に感染するなら、このような特定の記憶細胞が使われて即座に病原体は排除される。
リンパ球
獲得免疫に関与する細胞は特定の種類の白血球で、リンパ球と呼ばれている。その主要なタイプはB細胞とT細胞であり、骨髄の中の造血幹細胞に由来する[26]。B細胞は体液性免疫反応に関与し、T細胞は細胞性免疫応答に関与する。B細胞とT細胞は、特定の目標を認識する受容体分子をもっている。T細胞が病原体のような「異物」のターゲットを認識するには、抗原(病原体)が小片まで分解されて自己の受容体である主要組織適合遺伝子複合体(MHC、Major Histocompatibility Complex)分子と組み合わさって提示されねばならない。T細胞には細胞傷害性T細胞(キラーT細胞)とヘルパーT細胞の2種類の主要なサブタイプがある。細胞傷害性T細胞はMHCクラスI分子と結合した抗原のみを認識し、ヘルパーT細胞はMHCクラスII分子と結合した抗原のみを認識する。これらの2つの抗原提示の機構は、2タイプのT細胞の異なる役割を反映している。3番目のマイナーなサブタイプのT細胞としてγδT細胞があり、MHC受容体に結合しない、非加工の抗原を認識する[41]。
対照的に、B細胞の抗原に特有の受容体は、B細胞表面上の抗体分子であり、抗原加工なしに、病原体全体を認識する。B細胞上の抗体は、将来そのB細胞が産生する抗体のサンプルであるが多少の違いが存在する。B細胞の各々の増殖系は異なった抗体を発現し、B細胞の抗原受容体の完全な1セットは体が作ることができる全ての抗体を表すものである[26]。
細胞傷害性T細胞(CTL)

キラーT細胞が外来性ないし異常な抗原を表面にもった細胞に直接攻撃を加えている。[42]
細胞傷害性T細胞(CTL、キラーT細胞)はT細胞のサブグループで、ウイルス(および他の病原体)に感染した、損傷した、または機能不全の細胞を殺す[43]。
B細胞と同じく、各タイプのT細胞は異なる抗原を認識する。
細胞傷害性T細胞は、自身の持つT細胞受容体(TCR)が別の細胞のMHCクラスI受容体と複合体を作っている特定の抗原と結合するとき、活性化する。
このMHC-抗原複合体の認識は、T細胞上のCD8と呼ばれる共受容体によって助けられる。
それからこのT細胞は、このような抗原を保持したMHCクラスI受容体を発現させている細胞を捜して、体内をくまなく移動する。
活性化したT細胞がこのような細胞に接触すると、パーフォリンのような細胞傷害物質を放出する。パーフォリンは標的の細胞の細胞膜に孔を開け、イオン、水と毒素を侵入させる。
グラニュライシン(タンパク質分解酵素)と呼ばれるほかの毒性物質の侵入は、標的の細胞にアポトーシスを誘導する[44]。
T細胞による宿主細胞の殺害は、特にウイルスの複製を防ぐのに重要である。
T細胞の活性化は厳しく制御されていて、一般にきわめて強いMHC-抗原複合体の活性化シグナルか、ヘルパーT細胞による付加的な活性化シグナルを必要とする[44]。
ヘルパーT細胞 (Th)

ヘルパーT細胞の機能:抗原提示細胞(APC)はMHCクラスII分子(MHC2)上に抗原を提示する。ヘルパーT細胞はこれを認識し、これはCD4コレセプター(CD4+)の助けを得る。静止期ヘルパーT細胞の活性化によってサイトカインや他の刺激シグナル(緑の矢印)が放出され、膜、細胞傷害性T細胞、およびB細胞の活性を刺激する。B細胞への刺激は抗体産生につながる。B細胞とマクロファージへの刺激はヘルパーT細胞の増殖後に行われる。
ヘルパーT細胞(Th細胞)は自然免疫と獲得免疫の両方の免疫反応を調節していて、生体が特定の病原体に対して、どちらの免疫反応を行うか決定するのを助ける[45][46]。
ヘルパーT細胞には細胞を傷害する能力はなく、機能不全な細胞も殺さす、病原体も直接消さない。代わりに、他の免疫細胞への指示を司ることで免疫反応を統制している。
ヘルパーT細胞は、MHCクラスII分子と結合した抗原を認識するT細胞受容体(TCR)を発現している。そのMHCと抗原の複合体は、おなじくヘルパーT細胞のCD4共受容体によっても認識され、T細胞の活性化に作用するT細胞内の分子(例えばLck)を動員する。
ヘルパーT細胞のMHC:抗原複合体との関係は、細胞傷害性T細胞より弱い。
それは、細胞傷害性T細胞が1個のMHC:抗原複合体分子の交わりによって活性化するのに対し、ヘルパーT細胞の活性化には、多数(200〜300くらい)の受容体に、MHC:抗原複合体が付着しなければならない、ということである。
また、ヘルパーT細胞の活性化には、抗原提示細胞とより長い交わり時間を必要とする[47]。
休んでいたヘルパーT細胞は、活性化により、他の多くの細胞種の活性に影響するサイトカインを遊離する。
ヘルパーT細胞によって放出されるサイトカインのシグナルは、マクロファージの微生物殺滅作用と細胞傷害性T細胞や抗体を産生するB細胞の活動を強化する[3]。
加えて、ヘルパーT細胞の活性化は、CD40リガンド(別名CD154)のようなT細胞表面に発現している分子の調整量の上昇を引き起こす。
この分子は抗体産生B細胞を活性化するのに必要な代表的な付加的刺激シグナルとして働く[48]。
γδT細胞
γδT細胞はCD4+およびCD8+(αβ)T細胞とは対照的に別のT細胞受容体(TCR)をもち、ヘルパーT細胞、細胞傷害性T細胞、およびNK細胞と同じ性質を共有する。γδT細胞から応答を得る条件は完全には解明されていない。他のなじみのない変異型TCRをもったT細胞サブセット、例えばCD1d-拘束性ナチュラルキラーT細胞などと同様に、自然免疫と適応免疫の間を広くまたいでいる[49]。一方でγδT細胞は、この細胞はTCR遺伝子を再編成して受容体の多様性を生じること、そして記憶表現型も発達させることができることから、適応免疫の要素である。他方様々なサブセットは、制限されたTCRあるいはNK受容体が受容体のパターン認識に用いられることがあるため、自然免疫系の一部分をなす。例えばきわめて多数のヒトVγ9/Vδ2 T細胞は微生物によって産生される共通の分子に対して数時間以内に応答する。さらに高度に制限されたVδ1+
T細胞は上皮細胞が受けるストレスに応答するようだ[50]。
B細胞と抗体

抗体は2本の重鎖と2本の軽鎖から構成される。ユニークな可変部(可変領域)は対応する抗原を認識することが出来る。また、マクロファージは定常部に対する受容体を持っている[42]。
1個のB細胞は表面上の抗体が特定の外来抗原に結合すると病原体を認識することになる[51]。この抗原/抗体複合体はB細胞に取り込まれタンパク質分解プロセスによってペプチドにされる。B細胞は次にこれら抗原ペプチドを特異的なMHCクラスII分子上に提示する。MHCと抗原の複合体はその抗原と特異的に結合するヘルパーT細胞を引き寄せ、そのヘルパーT細胞がB細胞を活性化するリンフォカインを放出する[52]。B細胞が活性化されて増殖のための分裂を始めるとその子孫(形質細胞)はこの抗原を認識する特異的な抗体コピーを何百万分子も生産、分泌する。
これらの抗体は血管の血漿やリンパ管に入って循環する。抗体の実体は免疫グロブリンとよばれるタンパク質で、抗原を発現している細菌などの病原体に特異的に結合し、補体系の活性化あるいは食細胞による取り込みと破壊が起きるようマークを付ける。これをオプソニン化という。抗体は侵入病原体に対し、細菌の毒素に結合したりウイルスや細菌が細胞に感染する際に利用する受容体に妨害作用を及ぼして、直接中和することもできる[53]。
代替的適応免疫系
適応免疫の古典的な分子(例えば抗体やT細胞受容体)は顎をもった脊椎動物のみに存在するにも拘わらず、ヤツメウナギやメクラウナギのような原始的な無顎脊椎動物には独特なリンパ球由来の分子が発見されている。これらの動物には変異性リンパ球受容体(VLRs)と呼ばれる大きな一群の分子が備わり、顎をもった脊椎動物の抗原受容体のようにごくわずかな数(1つか2つ)の遺伝子のみから産生される。これらの分子は抗体と同じやり方で病原体の抗原に抗体と同じ程度の特異性をもって結合すると信じられている[54]。
免疫記憶
B細胞とT細胞が活性化されて複製を始めるとそれらの子孫細胞の中には長期間体内に残存する記憶細胞になるものがあるだろう。動物の生涯にわたってこれらの記憶細胞は各々の特異的な病原体に出合った記憶を保持し、病原体が再び感知されると強力な応答を発動できる。これは、個体の生涯にわたって病原体による感染に適応して起こり、免疫系が将来の接触に対して準備するものであるから、「適応」であると言える。免疫記憶は短期間の受動的な記憶の形か長期間にわたる能動的な記憶の形かのいずれかで成立しうる。
受動的な記憶
受動免疫 passive immunity は、抗体、細胞傷害性T細胞(CTL)といった既存の作用物質を投与して起こす免疫反応。
新生児はあらかじめ微生物に接触することはなく特に感染を受けやすい。そこで母親からいくつかの階層からなる受動防御が提供される。妊娠中抗体の特別の型IgGが胎盤を経由して直接母親から胎児に輸送される。したがってヒト新生児は誕生時すでに高レベルの母親と同じ抗原特異性の幅を持った抗体をもっている[55]。母乳も抗体をもっており赤ん坊の胃に移動し、新生児が自分自身の抗体を合成できるまで、細菌感染を防御する[56]。これは受動免疫であって、胎児は実際記憶細胞あるいは抗体を作らずそれらを母親から借用するだけであるから、この受動免疫は普通短期間のもので、数日から数カ月しか続かない。医学では、防御的な受動免疫が、ある個人から他人へ抗体リッチな血清を人工的に移すことでも行いうる[57]。
能動的な記憶と免疫処置

免疫応答が病原体の感染(あるいはワクチン初回投与)から始まり能動的な免疫記憶を形成して維持される時間的経過。
能動免疫 active immunity は、ワクチンなどの抗原を投与して誘導する免疫反応。
長期的な能動的な記憶は感染後B細胞およびT細胞の活性化によって獲得される。能動免疫は人工的にもワクチン接種によって成立させ得る。ワクチン接種(あるいは免疫処置と呼ばれる)の原理は病原体の抗原を導入し免疫系を刺激してその特定の病原体に対する特異的免疫を発達させその病原体由来の病気を起こさないようにすることである[3]。この意図的な免疫応答の誘導は免疫系が自然に作り出している特異性を利用していること、そうして免疫を誘導できるということによって成功している。ヒト集団の主要な死因の一つに感染症があることからワクチン処置は人類が発展させた免疫系の操作の中で最も効果のあるものである[58][26]。
大部分のウイルスワクチンは生きた弱毒化したウイルスをもとにしているが多くの細菌ワクチンは有害作用のない毒物質の成分など細菌の構成要素の非細胞成分をもとにしている[3]。多くの非細胞成分由来の抗原によるワクチンはあまり適応免疫応答を起こさないため、大部分の細菌ワクチンは、自然免疫の抗原提示細胞を活性化し免疫原性を最大にするアジュバントを添加して提供される[59]。
ヒトの免疫異常
免疫系は、特異性、誘導性、および適応性を取り込んできわめて効果的な構造をもつに至っている。しかし宿主防御に失敗することがあり、これは3つの大まかなカテゴリーに分けられる。免疫不全、自己免疫、過敏症、である。
免疫不全
免疫不全は免疫系の1つないしそれ以上の要素が機能しない場合に起きる。免疫系が病原体に対して応答する能力は、若くても年を取っても減退する。免疫応答は50歳位から免疫老化のために衰え始める[60][61]。先進国では肥満、アルコール依存症、薬物使用は免疫機能を弱める共通の原因である[61]。しかし開発途上国では栄養不良が免疫不全の最も多く見られる原因である[61]。十分なタンパク質を取らないダイエットは細胞性免疫や補体活性、食細胞機能、IgA抗体濃度、サイトカイン産生を損なう。栄養素であるイオン、銅、亜鉛、セレン、ビタミンA、C、E、B6、葉酸(ビタミンB9)が1つでも欠乏したら免疫応答は減退する[61]。加えて若いときに胸腺を遺伝的突然変異の原因か手術による摘出で失うと、重症の免疫不全を起こし、感染性が非常に高くなる[62]。
免疫不全は遺伝でも後天的でも生じうる[3]。慢性肉芽腫症では、食細胞の病原体破壊力が弱いということがあるが、遺伝性または先天性の免疫不全の例であるAIDSやいくつかのがんの型は、後天的な免疫不全を起こす[63][64]。
ある種のウイルスに感染することによって免疫機能が破壊され、様々な感染症・合併症を引き起こす病気が後天性免疫不全症候群(AIDS、エイズ)である。またこのウイルスをヒト免疫不全ウイルス (HIV) と呼ぶ。先天的に免疫機能が破綻しており、様々な感染症などを引き起こす病気はまとめて原発性免疫不全症候群と呼ばれる。
自己免疫
免疫応答の亢進は特に自己免疫病のような免疫不全の一方の極端をなす。ここでは免疫系は、自己と非自己を的確に区別できないで、自己の身体部分を攻撃する。普通の状態では多くのT細胞を抗体は自己のペプチドと反応する[65]。特別な細胞(胸腺および骨髄に潜む)の機能の1つに若いリンパ球に体内で産生されている自己抗原を提示し、自己抗原と認識した細胞を排除して自己免疫を防いでいる[51]。
過敏症
アレルギー(過敏症)は自己の組織に損傷を与える免疫応答である。クームス分類によると、5つの型に分けられる。
I型アレルギーは即時的な反応あるいはアナフィラキシー反応で、しばしばアレルギーに付随している。症状は穏やかな不快さから死に至るまで幅広い。I型アレルギーはマスト細胞や好塩基球が分泌するIgEが原因である[66]。
II型アレルギーは抗体が自己の細胞の抗原に結合してそれを破壊するようマークを付けることから起こる。これは抗体依存性(あるいは細胞傷害性)過敏症と呼ばれ、IgGやIgM抗体が原因である[66]。免疫複合体(抗原の凝集、補体タンパク質、およびIgGとIgM抗体)が様々な組織で沈着するとIII型アレルギーの反応が引き起こされる[66]。
IV型アレルギーは(細胞媒介性あるいは遅延型アレルギーとしても知られるが)生じるまでに普通は2〜3日かかる。IV型の反応は多くの自己免疫病や感染症で見られるが、接触皮膚炎(ツタウルシ)にも見られる場合がある。これらの反応に関与しているのはT細胞、単球およびマクロファージである[66]。
V型アレルギーは機序はII型と同様であるが、刺激性の部分だけが異なる。バセドウ病が代表的な疾患である。
他の機構
無脊椎動物はリンパ球を生み出していないし、抗体に基づいた体液性反応も生み出していないので多要素からなる適応免疫系は最初の脊椎動物に生じたと思われる[1]。しかし多くの種は脊椎動物の免疫のこれらの諸面の前駆機能として発現させている機構を活用している。免疫系は生物の機能構造としては最も簡単なものでさえあると見受けられる。細菌はバクテリオファージと呼ばれるウイルス病原体から守るために制限修飾系と呼ばれるユニークな防御機構を用いている[67]。原核生物も獲得免疫系をもっており、過去に接触したファージのゲノム断片を保持するのにCRISPR配列を用いてRNA干渉のような形でウイルスの複製を妨害することができる[68][69]。
パターン認識受容体は病原体に付随した分子を感知するのにほとんど総ての生物によって利用されている。ディフェンシンと呼ばれる抗微生物ペプチドは全ての動物および植物に見られる自然免疫応答の進化的に保存された要素の1つである[1]。補体系や食細胞も大部分の無脊椎動物で利用されている。リボヌクレアーゼとRNA干渉の反応経路は全ての真核生物で保存されていてウイルスに対する免疫応答に役割を果たしていると考えられる[70]。
動物と違い植物では食細胞を欠く。植物の大部分の免疫応答には植物から放出される全身的な化学的シグナルがある[71]。植物の一部が感染を受けるとその植物は局所的な過敏性の反応を起こす。そのことによって感染部位の細胞は速やかなアポトーシスを起こし他の植物への感染の広がりを阻止する。全身獲得抵抗性(SAR; Systemic Acquired Resistance)は防御反応の1つの型で、植物全体が特定の感染性病原体に抵抗するようにする[71]。RNAサイレンシング機構はウイルス複製をブロックできるのでこの全身的応答に特に重要である[72]。
腫瘍免疫

マクロファージががん細胞を認識したところ。左下~中央の仮足を持つ不定形の細胞(突起をたくさんもった大きな塊)が癌細胞、それに付着する凸凹の白い小さな球状細胞がマクロファージ。マクロファージは癌細胞と融合し、腫瘍細胞を殺す毒物を注入するだろう。がん治療における免疫療法は活発な医学研究領域の1つである[73]。
免疫系の他の重要な役割に腫瘍を見つけて排除することがある。腫瘍による形質転換細胞は正常細胞にない抗原を発現する。免疫系にとってこれらの抗原は非自己に見え、免疫細胞は形質転換した腫瘍細胞を攻撃する。腫瘍によって発現する抗原にはいくつかのソースがある。子宮頚がんを起こすヒトパピローマウイルスのような発がん性ウイルス由来のもの[74][75]。他は正常細胞では低レベルにしか見られないが腫瘍細胞で高レベルで見られるような自己タンパク質である。1例としてチロシナーゼと呼ばれる酵素があり、これは高レベルに発現するとある種の皮膚細胞(例えばメラノサイト)をメラノーマと呼ばれる腫瘍細胞に転換させる[76][77]。腫瘍抗原の3番目の可能なソースは正常細胞では細胞増殖や生存の制御に重要なタンパク質が普通に突然変異を起こしてがん誘起分子に変わる[74][78][79]。
免疫系の腫瘍に対する主な反応は異常細胞を細胞傷害性T細胞(CTL)やときにヘルパーT細胞の補助を受けて破壊することである[77][80]。腫瘍抗原はウイルス抗原と同じようにMHCクラスI分子上に提示される。これによって細胞傷害性T細胞(CTL)は腫瘍細胞を異常と見なす[81]。NK細胞も同じように腫瘍性の細胞を殺滅し特にMHCクラスI分子が正常に比べ少なく発現されている腫瘍細胞に対して作用する。このことは腫瘍細胞では一般的な現象として見られることである[82]。往々にして腫瘍細胞に対して抗体が産生され補体系にもそれらの細胞を破壊することが図られる[78]。
明らかに腫瘍の中には免疫系をうまく逃れてがんに向かうものがある[83]。腫瘍細胞はしばしば表面にMHCクラスI分子を発現する数が少ないので、細胞傷害性T細胞(CTL)による検出を免れる[81]。また腫瘍細胞の中には免疫応答を阻害する産物を放出するものがあり、例えばサイトカインTGF-βを分泌するとマクロファージやリンパ球の活性が抑制される[84]。加えて腫瘍細胞に対し免疫寛容が発達し免疫系が腫瘍細胞をもはや攻撃しないようにさせる場合もある[83]。
逆説的だが、腫瘍細胞がマクロファージをおびき寄せるサイトカインを放出し、マクロファージはその後腫瘍細胞が成長するようなサイトカインと増殖因子を産生するような場合、マクロファージは腫瘍の増殖を促進できる[85]。加えて腫瘍細胞における低酸素状態とマクロファージ産生のサイトカインの組合せは腫瘍細胞が転移をブロックするタンパク質を産生するのを減らし、がん細胞の広がりを助けることになる。
生理学的制御
ホルモンは免疫調節物質として働き、免疫系の感受性を変えることができる場合がある。例えば女性の性ホルモンは適応免疫応答に対しても[86] 自然免疫応答に対しても[87] 免疫賦活活性をもっていることが知られている。全身性エリテマトーデスのような自己免疫病は女性を選択的に襲うが、発症の時期はしばしば思春期であるという時期の一致がある。対照的にテストステロンのような男性ホルモンには免疫抑制力があるようだ[88]。他のホルモンにも免疫系を制御していると思われるものがあり、中でも有名なのがプロラクチン、成長ホルモン、ビタミンDである[89][90]。ホルモンレベルが年とともに減少を続けると、特に年老いた人々にとって免疫応答が減弱する原因となる[91]。反対にホルモンの中には免疫系の制御を受けるものがあり、目立つものとして、甲状腺ホルモンがあり、免疫系の制御を受ける[92]。
免疫系は睡眠や休息によって増強され[93]ストレスによって損なわれる[94]。
ダイエットは免疫系に影響することがある。例えば新鮮な果物、野菜、ある種の脂肪酸の豊富な食物は健康な免疫系を維持促進する[95]。同じように胎児の低栄養状態は免疫系に生涯続く損傷を与えうる[96]。伝統的な医学ではハーブの中に免疫系を刺激するものがあると信じられている。このようなハーブには、例えばエキナセア、甘草、距骨(玉縁)、サルビア、ニンニク、アメリカ・ニワトコの実、シイタケ、リンザイキノコ、ヒソップ、があり、さらにハチミツがある。研究によると、作用の仕方は複雑で特徴付けは困難にしても、そのようなハーブは実際免疫系を刺激することが示唆されている[97]。
医学における操作
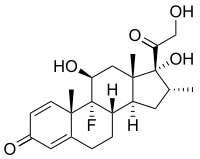
免疫抑制薬デキサメタゾン
免疫応答は、自己免疫、アレルギー、移植拒絶反応の結果起こる望まれない応答を抑制するよう、また、免疫系を大体逃れている病原体に対する防御反応を刺激するよう、操作しうる。ワクチン接種や血清療法は、免疫機構の抗原抗体反応を利用したものである。免疫抑制薬は自己免疫疾患や過剰な組織破壊が起こっている炎症の制御に、また器官移植後の移植拒絶を妨げるために使用される[26][98]。
抗炎症薬はしばしば炎症の影響を制御するのに用いられる。糖質コルチコイドはこれらの薬品の中で最も強力なものである。しかしこれらの薬品は多くの予想外の副作用をもちうる(例えば、中心性肥満、高血糖症、骨粗鬆症)。使用は厳重にコントロールしなければならない[99]。したがって、抗炎症薬は少量にしてメトトレキセートやアザチオプリンのような細胞傷害性あるいは免疫抑制薬との組合せで用いることがしばしば行われる。細胞傷害性の薬品は活性化T細胞のような分裂中の細胞を殺すような免疫応答を阻害する。しかし殺滅作用は区別できないから定常的に分裂している細胞とそれらの器官は影響を受け、これが毒性をもった副作用をもたらす[98]。シクロスポリンのような免疫抑制薬はシグナル伝達系を阻害することによってT細胞がシグナルに正しく反応するのを阻害する[100]。
大きな薬品(>500Da)は、特に繰り返し何度も投与されたり、投与量が大きいと、免疫応答を中和する場合がある。大きなペプチドおよびタンパク質(典型的には6,000Da以上)に基づいた薬品の効果には限界がある。薬品自身には免疫原性はなく、免疫原性のある物質と共投与される場合がある。このようなことはタキソールの場合時々見かける。ペプチドとタンパク質の免疫原性を予想するのにコンピュータによる方法が開発されて来ており特に治療用の抗体のデザイン、ウイルスのコート粒子に起こりそうな毒性突然変異を評価したり、ペプチドベースでの薬品処理の検証に有用である。初期のテクニックでは主にエピトープ域では親水性のアミノ酸が疎水性のアミノ酸より過剰に発現されているという観察に頼っていたが[101]、より最近の研究成果では、通常よく研究されたウイルスタンパク質に基づいてコンピュータはそれを学習材料として、知られたエピトープのデータベースに関しコンピュータが学習したテクニックに頼っている[102]。公開されてアクセスできるデータベースが、B細胞によって認識されるということが知られているエピトープのカタログ化を行うために確立されている[103]。免疫原性のバイオインフォマティクスに基づいた研究分野は新たに誕生したもので免疫インフォマティクスと言及される[104]。
病原体の操作
病原体の成功は宿主の免疫応答から逃れる能力に依存している。したがって病原体は宿主にうまく感染できるような方法を免疫を媒介にした破壊を免れつつ、いくつか発達させてきた[105]。細菌はしばしば物理的障壁についてはそれを分泌酵素で消化することによって切り抜ける。例えばII型分泌系の利用などである[106]。別の方法としてはIII型分泌系の利用があり、宿主細胞に穴を開ける管を挿入する。直接この管を通じて病原体から宿主へタンパク質を移動させる。管を通って輸送されるタンパク質はしばしば宿主防御を停止するのに用いられる[107]。
いくつかの病原体が自然免疫系から免れるのに用いている回避戦略は、細胞内複製である(細胞内病原性とも呼ばれる)。この場合病原体は生活史の大部分を宿主細胞内で過ごす。そこでは、免疫細胞、抗体、それに補体に直接接触することはなくそれらから保護される。細胞内病原体の例としてはウイルス、食中毒細菌のサルモネラ菌、真核生物の寄生虫であるマラリアを起こすもの(Plasmodium falciparum)やリーシュマニア症を起こすもの(Leishmania spp.)などである。結核菌(Mycobacterium tuberculosis)のような他の細菌は補体による溶解を阻止する保護カプセル中に生存する[108]。多くの病原体が宿主の免疫応答を弱め方向を間違うような化学物を分泌する[105]。細菌の中には免疫系の細胞やタンパク質から守るために生物的フィルムを形成するものがある。そのような生物的フィルムは多くの感染成功例に見られ、例えば嚢胞性線維症が特徴の慢性緑膿菌感染やバークホルデリア・セノセパシア感染がある[109]。ほかに抗体に結合する表面タンパク質を発現して抗体の効力を落とす細菌がある。この例には連鎖球菌(Gタンパク質)、黄色ブドウ球菌(Aタンパク質)、ペプトスプレプトコッカス・マグナス(Lタンパク質)がある[110]。
ウイルスが適応免疫系から免れる機構はもっと込み入っている。簡単な方法は、必須なエピトープは隠しもって全く必須でないウイルス表面上のエピトープを素早く変化させることである(アミノ酸か糖あるいは両方)。例えばHIVは、宿主のターゲット細胞に侵入するのに必須なウイルス外膜のタンパク質に絶えず突然変異を起こす。抗原のこれら頻繁な変化はこれらのタンパク質を対象とするワクチンを失敗させていることを説明するだろう。抗原を宿主分子でマスクする方法は宿主細胞から逃れるのによく見られる戦略である[111]。HIVではウイルスを覆う外膜は宿主細胞のもっとも外側の膜から作られている。このような"自己を覆い隠す"ウイルスは免疫系が"非自己"と認識するのを困難にしている[112]。
免疫学とその歴史
免疫学は免疫系の構造と機能を研究する科学である。
これは医学から生まれ初期の研究は病気に対する免疫の原因についてであった。免疫に最初に言及したのは、知られる限りでは、BC430年のアテネの悪疫流行の間である。ツキジデスは、以前病気にかかって回復した人々は患者を看護しても2度罹ることはないと記した[113]。このようにして観察された獲得免疫はのちにルイ・パスツールによって探求され、ワクチン接種の開発や病気の微生物原因論の提案に結びついた[114]。パスツールの理論は病気の当時流布していた瘴気論のような理論に真っ向から立ち向かうもので、この証明は、1891年にロバート・コッホによってなされた微生物が感染症の原因であることの証明まで待たねばならなかったが、コッホは1905年にノーベル賞に輝いた[115]。1901年のウォールター・リードによる、黄熱病ウイルス発見の際、ウイルスがヒト病原体として確認された[116]。
免疫学は19世紀終わりに向かって長足の進歩を遂げたが、急速な発展の中に体液性免疫および細胞性免疫の研究[117] で特に重要なのはパウル・エールリヒの仕事であり、彼は抗原-抗体反応の特異性の説明に側鎖説を唱えた。体液性免疫の理解に対する貢献は、細胞性免疫研究の立役者であるイリヤ・メチニコフと共同で1908年ノーベル賞受賞したことで認められた[118]。
脚注・出典
- ^ abc Beck, Gregory; Gail S. Habicht (1996年11月). “Immunity and the Invertebrates” (PDF). Scientific American: 60--66. http://www.scs.carleton.ca/~soma/biosec/readings/sharkimmu-sciam-Nov1996.pdf 2007年1月1日閲覧。.
^ Smith A.D. (Ed) Oxford dictionary of biochemistry and molecular biology. (1997) Oxford University Press. ISBN 0-19-854768-4- ^ abcdefghiAlberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?call=bv.View..ShowTOC&rid=mboc4.TOC&depth=2.
- ^ abLitman G, Cannon J, Dishaw L (2005年). “Reconstructing immune phylogeny: new perspectives”. Nat Rev Immunol 5 (11): 866--79. doi:10.1038/nri1712. PMID 16261174.
- ^ abcMayer, Gene (2006年). “Immunology - Chapter One: Innate (non-specific) Immunity”. Microbiology and Immunology On-Line Textbook. USC School of Medicine. 2007年1月1日閲覧。
^ Boyton R, Openshaw P (2002年). “Pulmonary defences to acute respiratory infection”. Br Med Bull 61: 1--12. doi:10.1093/bmb/61.1.1. PMID 11997295.
^ Agerberth B, Gudmundsson G. “Host antimicrobial defence peptides in human disease”. Curr Top Microbiol Immunol 306: 67--90. doi:10.1007/3-540-29916-5_3. PMID 16909918.
^ Moreau J, Girgis D, Hume E, Dajcs J, Austin M, O'Callaghan R (2001年). “Phospholipase A(2) in rabbit tears: a host defense against Staphylococcus aureus”. Invest Ophthalmol Vis Sci 42 (10): 2347--54. PMID 11527949. http://www.iovs.org/cgi/content/full/42/10/2347.
^ Hankiewicz J, Swierczek E (1974年). “Lysozyme in human body fluids”. Clin Chim Acta 57 (3): 205--9. doi:10.1016/0009-8981(74)90398-2. PMID 4434640.
^ Fair W, Couch J, Wehner N (1976年). “Prostatic antibacterial factor. Identity and significance”. Urology 7 (2): 169--77. doi:10.1016/0090-4295(76)90305-8. PMID 54972.
^ Yenugu S, Hamil K, Birse C, Ruben S, French F, Hall S (2003年). “Antibacterial properties of the sperm-binding proteins and peptides of human epididymis 2 (HE2) family; salt sensitivity, structural dependence and their interaction with outer and cytoplasmic membranes of Escherichia coli”. Biochem J 372 (Pt 2): 473--83. doi:10.1042/BJ20030225. PMID 12628001. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1223422&blobtype=pdf.
^ Gorbach S (1990年). “Lactic acid bacteria and human health”. Ann Med 22 (1): 37--41. doi:10.3109/07853899009147239. PMID 2109988.
^ Reid G, Bruce A (2003年). “Urogenital infections in women: can probiotics help?”. Postgrad Med J 79 (934): 428--32. doi:10.1136/pmj.79.934.428. PMID 12954951. http://pmj.bmj.com/cgi/content/full/79/934/428.
^ Salminen S, Gueimonde M, Isolauri E (2005年). “Probiotics that modify disease risk”. J Nutr 135 (5): 1294--8. PMID 15867327. http://jn.nutrition.org/cgi/content/full/135/5/1294.
^ Reid G, Jass J, Sebulsky M, McCormick J (2003年). “Potential uses of probiotics in clinical practice”. Clin Microbiol Rev 16 (4): 658--72. doi:10.1128/CMR.16.4.658-672.2003. PMID 14557292.
^ Hill L, Embil J (1986年). “Vaginitis: current microbiologic and clinical concepts”. CMAJ 134 (4): 321--31. PMID 3510698. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1490817&blobtype=pdf.
^ Medzhitov R (2007年). “Recognition of microorganisms and activation of the immune response”. Nature 449 (7164): 819--26. doi:10.1038/nature06246. PMID 17943118.
^ Matzinger P (2002年). “the danger model: a renewed sense of self”. Science 296 (5566): 301-5. doi:10.1126/science.1071059.
^ Kawai T, Akira S (2006年). “Innate immune recognition of viral infection”. Nat Immunol 7 (2): 131--7. doi:10.1038/ni1303. PMID 16424890.
^ Miller, SB (2006年). “Prostaglandins in Health and Disease: An Overview”. Seminars in Arthritis and Rheumatism 36 (1): 37--49. doi:10.1016/j.semarthrit.2006.03.005. PMID 16887467.
^ Ogawa Y, Calhoun WJ. (2006年). “The role of leukotrienes in airway inflammation”. J Allergy Clin Immunol. 118 (4): 789--98. doi:10.1016/j.jaci.2006.08.009. PMID 17030228.
^ Le Y, Zhou Y, Iribarren P, Wang J (2004年). “Chemokines and chemokine receptors: their manifold roles in homeostasis and disease”. Cell Mol Immunol 1 (2): 95--104. PMID 16212895. http://www.nature.com/bjp/journal/v147/n1s/pdf/0706475a.pdf.
^ Martin P, Leibovich S (2005年). “Inflammatory cells during wound repair: the good, the bad and the ugly”. Trends Cell Biol 15 (11): 599--607. doi:10.1016/j.tcb.2005.09.002. PMID 16202600.- ^ abRus H, Cudrici C, Niculescu F (2005年). “The role of the complement system in innate immunity”. Immunol Res 33 (2): 103--12. doi:10.1385/IR:33:2:103. PMID 16234578.
^ Mayer, Gene (2006年). “Immunology - Chapter Two: Complement”. Microbiology and Immunology On-Line Textbook. USC School of Medicine. 2007年1月1日閲覧。- ^ abcdefJaneway CA, Jr. et al (2005). Immunobiology. (6th ed. ed.). Garland Science. ISBN 0-443-07310-4.
^ Liszewski M, Farries T, Lublin D, Rooney I, Atkinson J (1996年). “Control of the complement system”. Adv Immunol 61: 201--83. doi:10.1016/S0065-2776(08)60868-8. PMID 8834497.
^ Sim R, Tsiftsoglou S (2004年). “Proteases of the complement system”. Biochem Soc Trans 32 (Pt 1): 21--7. doi:10.1042/BST0320021. PMID 14748705. http://www.biochemsoctrans.org/bst/032/0021/0320021.pdf.
^ Ryter A (1985年). “Relationship between ultrastructure and specific functions of macrophages”. Comp Immunol Microbiol Infect Dis 8 (2): 119--33. doi:10.1016/0147-9571(85)90039-6. PMID 3910340.
^ Langermans J, Hazenbos W, van Furth R (1994年). “Antimicrobial functions of mononuclear phagocytes”. J Immunol Methods 174 (1--2): 185--94. doi:10.1016/0022-1759(94)90021-3. PMID 8083520.
^ May R, Machesky L (2001年). “Phagocytosis and the actin cytoskeleton”. J Cell Sci 114 (Pt 6): 1061--77. PMID 11228151. http://jcs.biologists.org/cgi/reprint/114/6/1061.
^ Salzet M, Tasiemski A, Cooper E (2006年). “Innate immunity in lophotrochozoans: the annelids”. Curr Pharm Des 12 (24): 3043--50. doi:10.2174/138161206777947551. PMID 16918433.
^ Zen K, Parkos C (2003年). “Leukocyte-epithelial interactions”. Curr Opin Cell Biol 15 (5): 557--64. doi:10.1016/S0955-0674(03)00103-0. PMID 14519390.- ^ abStvrtinova', Viera; Ja'n Jakubovsky' and Ivan Huli'n (1995). Inflammation and Fever from Pathophysiology: Principles of Disease. Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. オリジナルの2001年7月11日時点によるアーカイブ。. https://web.archive.org/web/20010711220523/http://nic.savba.sk/logos/books/scientific/Inffever.html 2007年1月1日閲覧。.
^ Bowers, William (2006年). “Immunology -Chapter Thirteen: Immunoregulation”. Microbiology and Immunology On-Line Textbook. USC School of Medicine. 2007年1月4日閲覧。- ^ abGuermonprez P, Valladeau J, Zitvogel L, The'ry C, Amigorena S (2002年). “Antigen presentation and T cell stimulation by dendritic cells”. Annu Rev Immunol 20: 621--67. doi:10.1146/annurev.immunol.20.100301.064828. PMID 11861614.
^ Middleton D, Curran M, Maxwell L (2002年). “Natural killer cells and their receptors”. Transpl Immunol 10 (2--3): 147--64. doi:10.1016/S0966-3274(02)00062-X. PMID 12216946.
^ Kariyawasam H, Robinson D (2006年). “The eosinophil: the cell and its weapons, the cytokines, its locations”. Semin Respir Crit Care Med 27 (2): 117--27. doi:10.1055/s-2006-939514. PMID 16612762.
^ Krishnaswamy G, Ajitawi O, Chi D. “The human mast cell: an overview”. Methods Mol Biol 315: 13--34. PMID 16110146.
^ Pancer Z, Cooper M (2006年). “The evolution of adaptive immunity”. Annu Rev Immunol 24: 497--518. doi:10.1146/annurev.immunol.24.021605.090542. PMID 16551257.
^ Holtmeier W, Kabelitz D. “gammadelta T cells link innate and adaptive immune responses”. Chem Immunol Allergy 86: 151-83. PMID 15976493.- ^ ab“Understanding the Immune System: How it Works (PDF)”. National Institute of Allergy and Infectious Diseases (NIAID). 2007年1月3日時点のオリジナル[リンク切れ]よりアーカイブ。2007年1月1日閲覧。
^ Harty J, Tvinnereim A, White D (2000年). “CD8+ T cell effector mechanisms in resistance to infection”. Annu Rev Immunol 18: 275-308. doi:10.1146/annurev.immunol.18.1.275. PMID 10837060.- ^ abRadoja S, Frey A, Vukmanovic S (2006年). “T-cell receptor signaling events triggering granule exocytosis”. Crit Rev Immunol 26 (3): 265-90. PMID 16928189.
^ Abbas A, Murphy K, Sher A (1996年). “Functional diversity of helper T lymphocytes”. Nature 383 (6603): 787-93. doi:10.1038/383787a0. PMID 8893001.
^ McHeyzer-Williams L, Malherbe L, McHeyzer-Williams M (2006年). “Helper T cell-regulated B cell immunity”. Curr Top Microbiol Immunol 311: 59-83. doi:10.1007/3-540-32636-7_3. PMID 17048705.
^ Kovacs B, Maus M, Riley J, Derimanov G, Koretzky G, June C, Finkel T (2002年). “Human CD8+ T cells do not require the polarization of lipid rafts for activation and proliferation”. Proc Natl Acad Sci U S a 99 (23): 15006--11. doi:10.1073/pnas.232058599. PMID 12419850. http://www.ncbi.nlm.nih.gov/sites/entrez?db=pubmed&uid=12419850&cmd=showdetailview.
^ Grewal I, Flavell R (1998年). “CD40 and CD154 in cell-mediated immunity”. Annu Rev Immunol 16: 111-35. doi:10.1146/annurev.immunol.16.1.111. PMID 9597126.
^ Girardi M (2006年). “Immunosurveillance and immunoregulation by γδ T cells”. J Invest Dermatol 126 (1): 25--31. doi:10.1038/sj.jid.5700003. PMID 16417214.
^ Holtmeier W, Kabelitz D (2005年). “γδ T cells link innate and adaptive immune responses”. Chem Immunol Allergy 86: 151--183. doi:10.1159/000086659. PMID 15976493.- ^ abSproul T, Cheng P, Dykstra M, Pierce S (2000年). “A role for MHC class II antigen processing in B cell development”. Int Rev Immunol 19 (2--3): 139--55. doi:10.3109/08830180009088502. PMID 10763706.
^ Kehry M, Hodgkin P (1994年). “B-cell activation by helper T-cell membranes”. Crit Rev Immunol 14 (3--4): 221--38. PMID 7538767.
^ Bowers, William (2006年). “Immunology - Chapter nine: Cells involved in immune responses”. Microbiology and Immunology On-Line Textbook. USC School of Medicine. 2007年1月4日閲覧。
^ M.N. Alder, I.B. Rogozin, L.M. Iyer, G.V. Glazko, M.D. Cooper, Z. Pancer (2005年). “Diversity and Function of Adaptive Immune Receptors in a Jawless Vertebrate”. Science 310 (5756): 1970--1973. doi:10.1126/science.1119420. PMID 16373579.
^ Saji F, Samejima Y, Kamiura S, Koyama M (1999年). “Dynamics of immunoglobulins at the feto-maternal interface”. Rev Reprod 4 (2): 81--9. doi:10.1530/ror.0.0040081. PMID 10357095. http://ror.reproduction-online.org/cgi/reprint/4/2/81.pdf.
^ Van de Perre P (2003年). “Transfer of antibody via mother's milk”. Vaccine 21 (24): 3374--6. doi:10.1016/S0264-410X(03)00336-0. PMID 12850343.
^ Keller, Margaret A. and E. Richard Stiehm (2000年). “Passive Immunity in Prevention and Treatment of Infectious Diseases”. Clinical Microbiology Reviews 13 (4): 602--614. doi:10.1128/CMR.13.4.602-614.2000. PMID 11023960. http://cmr.asm.org/cgi/content/full/13/4/602.
^ Death and DALY estimates for 2002 by cause for WHO Member States. World Health Organization. Retrieved on 2007-01-01.
^ Singh M, O'Hagan D (1999年). “Advances in vaccine adjuvants”. Nat Biotechnol 17 (11): 1075--81. doi:10.1038/15058. PMID 10545912.
^ Aw D, Silva A, Palmer D (2007年). “Immunosenescence: emerging challenges for an ageing population”. Immunology 120 (4): 435--446. doi:10.1111/j.1365-2567.2007.02555.x. PMID 17313487.- ^ abcdChandra, RK (1997年). “Nutrition and the immune system: an introduction”. American Journal of Clinical Nutrition Vol 66: 460S--463S. PMID 9250133. http://www.ajcn.org/cgi/content/abstract/66/2/460S. Free full-text pdf available
^ Miller JF (2002年). “The discovery of thymus function and of thymus-derived lymphocytes”. Immunol. Rev. 185: 7--14. PMID 12190917. http://www.blackwell-synergy.com/doi/abs/10.1034/j.1600-065X.2002.18502.x.
^ Joos L, Tamm M (2005年). “Breakdown of pulmonary host defense in the immunocompromised host: cancer chemotherapy”. Proc Am Thorac Soc 2 (5): 445--8. doi:10.1513/pats.200508-097JS. PMID 16322598. http://pats.atsjournals.org/cgi/content/full/2/5/445.
^ Copeland K, Heeney J (1996年). “T helper cell activation and human retroviral pathogenesis”. Microbiol Rev 60 (4): 722--42. PMID 8987361. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=239461&blobtype=pdf.
^ Miller J (1993年). “Self-nonself discrimination and tolerance in T and B lymphocytes”. Immunol Res 12 (2): 115--30. doi:10.1007/BF02918299. PMID 8254222.- ^ abcdGhaffar, Abdul (2006年). “Immunology - Chapter Seventeen: Hypersensitivity Reactions”. Microbiology and Immunology On-Line Textbook. USC School of Medicine. 2007年1月1日閲覧。
^ Bickle T, Kru"ger D (1993年). “Biology of DNA restriction”. Microbiol Rev 57 (2): 434--50. PMID 8336674. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=372918&blobtype=pdf.
^ Barrangou R, Fremaux C, Deveau H, et al (2007年3月). “CRISPR provides acquired resistance against viruses in prokaryotes”. Science (journal) 315 (5819): 1709--12. doi:10.1126/science.1138140. PMID 17379808.
^ Brouns SJ, Jore MM, Lundgren M, et al (2008年8月). “Small CRISPR RNAs guide antiviral defense in prokaryotes”. Science (journal) 321 (5891): 960--4. doi:10.1126/science.1159689. PMID 18703739.
^ Stram Y, Kuzntzova L. (2006年). “Inhibition of viruses by RNA interference”. Virus Genes 32 (3): 299--306. doi:10.1007/s11262-005-6914-0. PMID 16732482.- ^ abSchneider, David (Spring 2005). “Innate Immunity - Lecture 4: Plant immune responses”. Stanford University Department of Microbiology and Immunology. 2007年6月9日時点のオリジナル[リンク切れ]よりアーカイブ。2007年1月1日閲覧。
^ Baulcombe D (2004年). “RNA silencing in plants”. Nature 431 (7006): 356--63. doi:10.1038/nature02874. PMID 15372043.
^ Morgan R et al. (2006年). “Cancer regression in patients after transfer of genetically engineered lymphocytes”. Science 314: 126--129. doi:10.1126/science.1129003. PMID 16946036.- ^ abAndersen MH, Schrama D, Thor Straten P, Becker JC (2006年). “Cytotoxic T cells”. J Invest Dermatol 126 (1): 32--41. doi:10.1038/sj.jid.5700001. PMID 16417215.
^ Boon T, van der Bruggen P (1996年). “Human tumor antigens recognized by T lymphocytes”. J Exp Med 183: 725--29. doi:10.1084/jem.183.3.725. PMID 8642276.
^ Castelli C, Rivoltini L, Andreola G, Carrabba M, Renkvist N, Parmiani G (2000年). “T cell recognition of melanoma-associated antigens”. J Cell Physiol 182: 323--31. doi:10.1002/(SICI)1097-4652(200003)182:3<323::AID-JCP2>3.0.CO;2-#. PMID 10653598.- ^ abRomero P, Cerottini JC, Speiser DE (2006年). “The human T cell response to melanoma antigens”. Adv Immunol. 92: 187--224. doi:10.1016/S0065-2776(06)92005-7. PMID 17145305.
- ^ abGuevara-Patino JA, Turk MJ, Wolchok JD, Houghton AN (2003年). “Immunity to cancer through immune recognition of altered self: studies with melanoma”. Adv Cancer Res. 90: 157--77. doi:10.1016/S0065-230X(03)90005-4. PMID 14710950.
^ Renkvist N, Castelli C, Robbins PF, Parmiani G (2001年). “A listing of human tumor antigens recognized by T cells”. Cancer Immunol Immunother 50: 3--15. doi:10.1007/s002620000169. PMID 11315507.
^ Gerloni M, Zanetti M. (2005年). “CD4 T cells in tumor immunity”. . Springer Semin Immunopathol 27 (1): 37--48. doi:10.1007/s00281-004-0193-z. PMID 15965712.- ^ abSeliger B, Ritz U, Ferrone S (2006年). “Molecular mechanisms of HLA class I antigen abnormalities following viral infection and transformation”. Int J Cancer 118 (1): 129--38. doi:10.1002/ijc.21312. PMID 16003759.
^ Hayakawa Y, Smyth MJ. (2006年). “Innate immune recognition and suppression of tumors”. Adv Cancer Res 95: 293--322. doi:10.1016/S0065-230X(06)95008-8. PMID 16860661.- ^ abSeliger B (2005年). “Strategies of tumor immune evasion”. BioDrugs 19 (6): 347--54. doi:10.2165/00063030-200519060-00002. PMID 16392887.
^ Frumento G, Piazza T, Di Carlo E, Ferrini S (2006年). “Targeting tumor-related immunosuppression for cancer immunotherapy”. Endocr Metab Immune Disord Drug Targets 6 (3): 233--7. doi:10.2174/187153006778250019. PMID 17017974.
^ Stix, Gary (2007年7月). “A Malignant Flame” (PDF). Scientific American: 60--67. オリジナルの2011年7月16日時点によるアーカイブ。. https://web.archive.org/web/20110716015048/http://podcast.sciam.com/daily/pdf/sa_d_podcast_070619.pdf 2007年1月1日閲覧。.
^ Wira, CR; Crane-Godreau M, Grant K (2004). “Endocrine regulation of the mucosal immune system in the female reproductive tract”. In In: Ogra PL, Mestecky J, Lamm ME, Strober W, McGhee JR, Bienenstock J (eds.). Mucosal Immunology. San Francisco: Elsevier. ISBN 0124915434.
^ Lang, TJ (2004年). “Estrogen as an immunomodulator”. Clin Immunol 113: 224–230. doi:10.1016/j.clim.2004.05.011. PMID 15507385.
Moriyama, A; Shimoya K, Ogata I et al. (1999年). “Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle”. Molecular Human Reproduction 5: 656–661. doi:10.1093/molehr/5.7.656. PMID 10381821. http://molehr.oxfordjournals.org/cgi/content/full/5/7/656.
Cutolo, M; Sulli A, Capellino S, Villaggio B, Montagna P, Seriolo B, Straub RH (2004年). “Sex hormones influence on the immune system: basic and clinical aspects in autoimmunity”. Lupus 13: 635–638. doi:10.1191/0961203304lu1094oa. PMID 15485092.
King, AE; Critchley HOD, Kelly RW (2000年). “Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial role”. Molecular Human Reproduction 6: 191–196. doi:10.1093/molehr/6.2.191. PMID 10655462. http://molehr.oxfordjournals.org/cgi/content/full/6/2/191.
^ Fimmel; Zouboulis CC (2005年). “Influence of physiological androgen levels on wound healing and immune status in men”. Aging Male 8: 166–174. doi:10.1080/13685530500233847. PMID 16390741.
^ Dorshkind, K; Horseman ND (2000年). “The Roles of Prolactin, Growth Hormone, Insulin-Like Growth Factor-I, and Thyroid Hormones in Lymphocyte Development and Function: Insights from Genetic Models of Hormones and Hormone Receptor Deficiency”. Endocrine Reviews 21: 292–312. doi:10.1210/er.21.3.292. PMID 10857555. http://edrv.endojournals.org/cgi/content/full/21/3/292?maxtoshow=&HITS=10&hits=10&RESULTFORMAT=&author1=Dorshkind%2C+K%3B+Horseman+ND+&searchid=1&FIRSTINDEX=0&sortspec=relevance&resourcetype=HWCIT.
^ Nagpal, Sunil; Songqing Naand and Radhakrishnan Rathnachalam (2005年). “Noncalcemic Actions of Vitamin D Receptor Ligands”. Endocrine Reviews 26 (5): 662–687. doi:10.1210/er.2004-0002. PMID 15798098. http://edrv.endojournals.org/cgi/content/full/26/5/662. .
^ Hertoghe, T (2005年). “The “multiple hormone deficiency” theory of aging: Is human senescence caused mainly by multiple hormone deficiencies?”. Annals of the New York Academy of Science 1051: 448–465. doi:10.1196/annals.1322.035. PMID 16399912.
^ Klein, JR (2006年). “The immune system as a regulator of thyroid hormone activity”. Exp Biol Med 231: 229–236. PMID 16514168.
^ Lange, T; Perras B, Fehm HL, Born J (2003年). “Sleep Enhances the Human Antibody response to Hepatitis A Vaccination”. Psychosomatic Medicine 65: 831–835. doi:10.1097/01.PSY.0000091382.61178.F1. PMID 14508028. http://www.psychosomaticmedicine.org/cgi/content/full/65/5/831.
^ Khansari, DN; Murgo AJ, Faith RE (1990年). “Effects of stress on the immune system”. Immunology Today 11: 170–175. doi:10.1016/0167-5699(90)90069-L. PMID 2186751.
^ Pond, CM (2005年). “Adipose tissue and the immune system”. Prostaglandins, Leukotrienes, and Essential Fatty Acids 73: 17--30. doi:10.1016/j.plefa.2005.04.005. PMID 15946832.
^ Langley-Evans, SC; Carrington LJ (2006年). “Diet and the developing immune system”. Lupus 15: 746–752. doi:10.1177/0961203306070001. PMID 17153845.
^ Spelman, K; Burns J, Nichols D, Winters N, Ottersberg S, Tenborg M (2006年). “Modulation of cytokine expression by traditional medicines: a review of herbal immunomodulators”. Alternative Medicine reviews 11: 128–150. PMID 16813462.
Brush, J; Mendenhall E, Guggenheim A, Chan T, Connelly E, Soumyanth A, Buresh R, Barrett R, Zwickey H (2006年). “The effect of Echinacea purpurea, Astragalus membranaceus and Glycyrrhiza glabra on CD69 expression and immune cell activation in humans”. Phytotherapy Research 20: 687–695. doi:10.1002/ptr.1938. PMID 16807880.- ^ abTaylor A, Watson C, Bradley J (2005年). “Immunosuppressive agents in solid organ transplantation: Mechanisms of action and therapeutic efficacy”. Crit Rev Oncol Hematol 56 (1): 23--46. doi:10.1016/j.critrevonc.2005.03.012. PMID 16039869.
^ Barnes P (2006年). “Corticosteroids: the drugs to beat”. Eur J Pharmacol 533 (1--3): 2--14. doi:10.1016/j.ejphar.2005.12.052. PMID 16436275.
^ Masri M (2003年). “The mosaic of immunosuppressive drugs”. Mol Immunol 39 (17--18): 1073--7. doi:10.1016/S0161-5890(03)00075-0. PMID 12835079.
^ Welling GW, Wiejer WJ, van der Zee R, Welling-Werster S. (1985年). “Prediction of sequential antigenic regions in proteins”. J Mol Recognit 88 (2): 215--8. PMID 2411595.
^ Sollner J, Mayer B. (2006年). “Machine learning approaches for prediction of linear B-cell epitopes on proteins”. Journal of Molecular Recognition 19 (3): 200--8. doi:10.1002/jmr.771. PMID 16598694.
^ Saha S, Bhasin M, Raghava GP. (2005年). “Bcipep: a database of B-cell epitopes”. BMC Bioinformatics 6 (1): 79. doi:10.1186/1471-2105-6-79. PMID 15921533.
^ Flower DR, Doytchinova IA. (2002年). “Immunoinformatics and the prediction of immunogenicity”. Appl Bioinformatics 1 (4): 167--76. PMID 15130835.- ^ abFinlay B, McFadden G (2006年). “Anti-immunology: evasion of the host immune system by bacterial and viral pathogens”. Cell 124 (4): 767--82. doi:10.1016/j.cell.2006.01.034. PMID 16497587.
^ Cianciotto NP. (2005年). “Type II secretion: a protein secretion system for all seasons”. Trends Microbiol. 13 (12): 581--8. doi:10.1016/j.tim.2005.09.005. PMID 16216510.
^ Winstanley C, Hart CA (2001年). “Type III secretion systems and pathogenicity islands”. J Med Microbiol. 50 (2): 116--26. PMID 11211218.
^ Finlay B, Falkow S (1997年). “Common themes in microbial pathogenicity revisited”. Microbiol Mol Biol Rev 61 (2): 136--69. PMID 9184008. http://mmbr.asm.org/cgi/reprint/61/2/136.pdf.
^ Kobayashi H (2005年). “Airway biofilms: implications for pathogenesis and therapy of respiratory tract infections”. Treat Respir Med 4 (4): 241--53. doi:10.2165/00151829-200504040-00003. PMID 16086598.
^ Housden N, Harrison S, Roberts S, Beckingham J, Graille M, Stura E, Gore M (2003年). “Immunoglobulin-binding domains: Protein L from Peptostreptococcus magnus”. Biochem Soc Trans 31 (Pt 3): 716--8. doi:10.1042/BST0310716. PMID 12773190. http://www.biochemsoctrans.org/bst/031/0716/0310716.pdf.
^ Burton, Dennis R.; Robyn L. Stanfield and Ian A. Wilson (2005年). “Antibody vs. HIV in a clash of evolutionary titans”. Proc Natl Acad Sci U S A. 102 (42): 14943--8. doi:10.1073/pnas.0505126102. PMID 16219699.
^ Cantin R, Methot S, Tremblay MJ. (2005年). “Plunder and stowaways: incorporation of cellular proteins by enveloped viruses”. J Virol. 79 (11): 6577-87. doi:10.1128/JVI.79.11.6577-6587.2005. PMID 15890896.
^ Retief F, Cilliers L (1998年). “The epidemic of Athens, 430-426 BC”. S Afr Med J 88 (1): 50--3. PMID 9539938.
^ Plotkin S (2005年). “Vaccines: past, present and future”. Nat Med 11 (4 Suppl): S5--11. doi:10.1038/nm1209. PMID 15812490.
^ The Nobel Prize in Physiology or Medicine 1905 Nobelprize.org Accessed January 8 2007.
^ Major Walter Reed, Medical Corps, U.S. Army Archived 2007年10月23日, at the Wayback Machine. Walter Reed Army Medical Center. Accessed January 8 2007.
^
Metchnikoff, Elie; Translated by F.G. Binnie. (1905) (Full Text Version: Google Books). Immunity in Infective Diseases. Cambridge University Press. http://books.google.com/books?vid=OCLC03666307&id=ywKp9YhK5t0C&printsec=titlepage&vq=Ehrlich&dq=history+of+humoral+immunity.
^ The Nobel Prize in Physiology or Medicine 1908 Nobelprize.org Accessed January 8 2007
関連項目
関連項目が多すぎます。関連の深い項目だけに絞ってください。必要ならば一覧記事として独立させることも検討してください。(2016年9月) |
- 免疫
- 細胞性免疫
液性免疫(体液性免疫)
細胞(造血幹細胞由来)
白血球- 顆粒球:好中球、好酸球、好塩基球
- リンパ球
B細胞:形質細胞- T細胞
ナチュラルキラー細胞(NK細胞)
- 単球:マクロファージ(大食細胞)
- 顆粒球:好中球、好酸球、好塩基球
樹状細胞:ランゲルハンス細胞
肥満細胞(マスト細胞)
- 分子
補体系:補体結合反応- サイトカイン
- ヒスタミン
- 異物
病原体:感染、感染症、日和見感染
腫瘍:腫瘍免疫
臓器移植:拒絶反応
- 炎症
- 異常、障害、免疫疾患
アレルギー:アナフィラキシー
自己抗体:自己免疫疾患(自律免疫疾患)
- 医薬品
- 免疫抑制剤
- 抗炎症薬
ワクチン:トキソイド(類毒素)
外部リンク
Immunity In Depth(英語) - (オレゴン州大学・ライナス・ポーリング研究所)
血液、免疫 (英語)- 「ここまで分かった亜鉛の免疫システムにおける役割」『日本衛生学雑誌』Vol.68 (2013) No.3 p.145-152
| ||||||||||||||


