Mushroom bodies

Mushroom bodies visible in a Drosophila brain as two stalks. From Jenett et al., 2006[1]
The mushroom bodies or corpora pedunculata are a pair of structures in the brain of insects, other arthropods, and some annelids (notably the ragworm Platynereis dumerilii).[2] They are also known to play a role in olfactory learning and memory. In most insects, the mushroom bodies and the lateral horn are the two higher brain regions that receive olfactory information from the antennal lobe via projection neurons.[3]
Contents
1 Structure
2 Function
3 Drosophila melanogaster
3.1 Cellular memory traces
3.2 cAMP dynamics
3.3 PKA dynamics
4 See also
5 References
6 Further reading
7 External links
Structure
Mushroom bodies are usually described as neuropils, i.e. as dense networks of neuronal processes (dendrite and axon terminals) and glia. They get their name from their roughly hemispherical calyx, a protuberance that is joining to the rest of the brain by a central nerve tract or peduncle. They were first identified in 1850[4] by the French biologist Félix Dujardin.[5]
Most of our current knowledge of the mushroom bodies comes from studies of a few species of insect, especially the cockroach Periplaneta americana, the honey bee Apis mellifera, the locust and the fruit fly Drosophila melanogaster. Studies of fruit fly mushroom bodies have been particularly important for understanding the genetic basis of mushroom body functioning, since their genome has been sequenced and a vast number of tools to manipulate their gene expression exist.
In the insect brain, the peduncles of the mushroom bodies extend through the midbrain. They are mainly composed of the long, densely packed nerve fibres of the Kenyon cells, the intrinsic neurons of the mushroom bodies. These cells have been found in the mushroom bodies of all species that have been investigated, though their number varies; for example fruit flies have around 2,500 whereas cockroaches have about 200,000.
Function
Mushroom bodies are largest in the Hymenoptera, which are known to have particularly elaborate control over olfactory behaviours. However, since mushroom bodies are found in anosmic primitive insects, their role is likely to extend beyond olfactory processing. Anatomical studies suggest a role in the processing of visual and mechanosensory input in some species.[6] In Hymenoptera in particular, subregions of the mushroom body neuropil are specialized to receive olfactory, visual, or both types of sensory input.[7] In Hymenoptera, olfactory input is layered in the calyx. In ants, several layers can be discriminated that correspond to different clusters of glomeruli in the antennal lobes, perhaps corresponding to different classes of odors.[3][8] There are two main groups of projection neurons that divide the antennal lobe into two main regions, an anterior and a posterior. Projection neuron groups are segregated innervating glomeruli groups separately and sending axons by separate routes, either through the medial-antenno protocerebral tract (m-APT) or through the lateral-anteno protocerebral tract (l-APT), and connecting with two layers in the calyx of the mushroom bodies. In these layers is represented topographically the organization of the two efferent regions of the antennal lobe, establishing a coarse odotopic map of the antennal lobe in the region of the lip of the mushroom bodies.[3][8]
Mushroom bodies are known to be involved in learning and memory, particularly for smell, and thus are the subject of current intense research. In larger insects, studies suggest that mushroom bodies have other learning and memory functions, like associative memory, sensory filtering, motor control, and place memory. Research implies that mushroom bodies generally act as a sort of coincidence detector, integrating multimodal inputs[3] and creating novel associations, thus illuminating their role in learning and memory.[9]
Information about odors may be encoded in the mushroom body by the identities of the responsive neurons as well as the timing of their spikes.[10] Experiments in locusts have shown that Kenyon cells have their activity synchronized to 20-Hz neural oscillations and are particularly responsive to projection neuron spikes at specific phases of the oscillatory cycle.[11]
Drosophila melanogaster

Mushroom body lobes of D. melanogaster, with α/β, α’/β’, and γ neurons visible. From Davis, 2011[12]
We know that mushroom body structures are important for olfactory learning and memory in Drosophila because their ablation destroys this function.[13] The mushroom body is also able to combine information from the internal state of the body and the olfactory input to determine innate behavior.[14] The exact roles of the specific neurons making up the mushroom bodies are still unclear. However, these structures are studied extensively because much is known about their genetic make-up. There are three specific classes of neurons that make up the mushroom body lobes: α/β, α’/β’, and γ neurons, which all have distinct gene expression. A topic of current research is which of these substructures in the mushroom body are involved in each phase and process of learning and memory.[15]Drosophila mushroom bodies are also often used to study learning and memory and are manipulated due to their relatively discrete nature. Typically, olfactory learning assays consist of exposing flies to two odors separately; one is paired with electric shock pulses (the conditioned stimulus, or CS+), and the second is not (unconditioned stimulus, or US). After this training period, flies are placed in a T-maze with the two odors placed individually on either end of the horizontal ‘T’ arms. The percent of flies that avoid the CS+ is calculated, with high avoidance considered evidence of learning and memory.[16]
Cellular memory traces
Recent studies combining odor conditioning and cellular imaging have identified six memory traces that coincide with molecular changes in the Drosophila olfactory system. Three of these traces are associated with early forming behavioral memory. One such trace was visualized in the antennal lobe (AL) by synapto-pHluorin reporter molecules. Immediately after conditioning, an additional set of projection neurons in a set of eight glomeruli in the AL becomes synaptically activated by the conditioned odor, and lasts for only 7 minutes.[17] A second trace is detectable by GCaMP expression, and thus an increase in Ca2+ influx, in the α’/β’ axons of the mushroom body neurons.[18] This is a longer-lasting trace, present for up to one hour following conditioning. The third memory trace is the reduction of activity of the anterior-paired lateral neuron, which acts as a memory formation suppressor through one of its inhibitory GABAergic receptors. Decrease in calcium response of APL neurons and subsequent decrease in GABA release onto the mushroom bodies persisted up to 5 minutes after odor conditioning.[19]
The intermediate term memory trace is dependent on expression of the amn gene located in dorsal paired medial neurons. An increase in calcium influx and synaptic release that innervates the mushroom bodies becomes detectable approximately 30 minutes after pairing of electric shock with an odor, and persists for at least an hour.[20] Both long-term memory traces that have been mapped depend on activity and protein synthesis of CREB and CaMKII, and only exist after spaced conditioning. The first trace is detected in α/β neurons between 9 and 24 hours after conditioning, and is characterized by an increase in calcium influx in response to the conditioned odor.[21] The second long-term memory trace forms in the γ mushroom bodies and is detected by increase calcium influx between 18 and 24 hours after conditioning [22]
cAMP dynamics
Cyclic adenosine monophosphate (cAMP or cyclic AMP) is a second messenger that has been implicated in facilitating mushroom body calcium influx in Drosophila melanogaster MBNs. cAMP elevation induces presynaptic plasticity in Drosophila. cAMP levels are affected by both neurotransmitters, such as dopamine and octopamine, and odors themselves. Dopamine and octopamine are released by mushroom body interneurons, while odors directly activate neurons in the olfactory pathway, causing calcium influx through voltage-gated calcium channels.[23]
In a classical conditioning paradigm, pairing neuronal depolarization (via acetylcholine application to represent the odor or CS) with subsequent dopamine application (to represent the shock or US), results in a synergistic increase in cAMP in the mushroom body lobes.[23] These results suggest that the mushroom body lobes are a critical site of CS/US integration via the action of cAMP. This synergistic effect was originally observed in Aplysia, where pairing calcium influx with activation of G protein signaling by serotonin generates a similar synergistic increase in cAMP.[24]
Additionally, this synergistic increase in cAMP is mediated by and dependent on rutabaga adenylyl cyclase (rut AC), which is sensitive to both calcium (which results from voltage-gated calcium channel opening by odors) and G protein stimulation (caused by dopamine).[23] While a forward pairing of neuronal depolarization and dopamine, (acetylcholine followed by dopamine) results in a synergistic increase in cAMP, a forward pairing of neuronal depolarization and octopamine produces a sub-additive effect on cAMP.[23] More specifically, this means that this pairing produces significantly less cAMP than the sum of each stimulus individually in the lobes. Therefore, rut AC in mushroom body neurons works as a coincidence detector with dopamine and octopamine functioning bidirectionally to affect cAMP levels.[23]
PKA dynamics
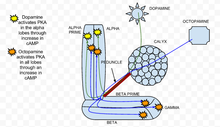
Spatial regulation of PKA dynamics in Drosophila mushroom body.
PKA, also known as protein kinase A, has been found to play an important role in learning and memory in Drosophila.[25] When calcium enters a cell and binds with calmodulin, it stimulates adenylate cyclase (AC), which is encoded by the rutabaga gene (rut).[26] This AC activation increases the concentration of cAMP, which activates PKA.[26] When dopamine, an aversive olfactory stimulant, is applied it activates PKA specifically in the vertical mushroom body lobes.[26] This spatial specificity is regulated by the dunce (dnc) PDE, a cAMP-specific phosphodiesterase. If the dunce gene is abolished, as found in the dnc mutant, the spatial specificity is not maintained. In contrast, an appetitive stimulation created by an octopamine application increases PKA in all lobes.[26] In the rut mutant, a genotype in which the rutabaga is abolished, the responses to both dopamine and octopamine were greatly reduced and close to experimental noise.
Acetylcholine, which represents the conditioned stimulus, leads to a strong increase in PKA activation compared to stimulation with dopamine or octopamine alone.[26] This reaction is abolished in rut mutants, which demonstrates that PKA is essential for sensory integration.[26] The specificity of activation of the alpha lobe in the presence of dopamine is maintained when dopamine is in combination with acetylcholine.[26] Essentially, during a conditioning paradigm when a conditioned stimulus is paired with an unconditioned stimulus, PKA exhibits heightened activation. This shows that PKA is required for conditioned learning in Drosophila melanogaster.
See also
- Cerebral cortex
References
^ Jenett A.; Schindelin J. E.; Heisenberg M. (2006). "The Virtual Insect Brain protocol: creating and comparing standardized neuroanatomy". BMC Bioinformatics. 7: 544. doi:10.1186/1471-2105-7-544. PMC 1769402. PMID 17196102..mw-parser-output cite.citationfont-style:inherit.mw-parser-output qquotes:"""""""'""'".mw-parser-output code.cs1-codecolor:inherit;background:inherit;border:inherit;padding:inherit.mw-parser-output .cs1-lock-free abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-subscription abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registrationcolor:#555.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration spanborder-bottom:1px dotted;cursor:help.mw-parser-output .cs1-hidden-errordisplay:none;font-size:100%.mw-parser-output .cs1-visible-errorfont-size:100%.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-formatfont-size:95%.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-leftpadding-left:0.2em.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-rightpadding-right:0.2em
^ ^ Tomer, R.; Denes, A. S.; Tessmar-Raible, K.; Arendt, D. (2010). "Profiling by image registration reveals common origin of annelid mushroom bodies and vertebrate pallium". Cell 142 (5): 800–809. doi:10.1016/j.cell.2010.07.043.
PMID 20813265
^ abcd Gronenberg, W.; López-Riquelme, G.O. (February 2014). "Multisensory convergence in the mushroom bodies of ants and bees". Acta Biologica Hungarica. 55 (1–4): 31–37. doi:10.1556/ABiol.55.2004.1-4.5.
^ Strausfeld N. J.; Hansen L; Li Y; Gomez R. S.; Ito K. (1998). "Evolution, discovery, and interpretations of arthropod mushroom bodies". Learn. Mem. 5 (1–2): 11–37. doi:10.1101/lm.5.1.11. PMC 311242. PMID 10454370.
^ Dujardin, F. 1850. Mémoire sur le système nerveux des insectes. Ann. Sci. Nat. Zool. 14: 195-206.
^ Zars, Troy (December 2000). "Behavioral functions of the insect mushroom bodies". Curr Opin Neurobiol. 10 (6): 790–5. doi:10.1016/S0959-4388(00)00147-1.
^ Mobbs, P. G. (1982). "The Brain of the Honeybee Apis Mellifera. I. The Connections and Spatial Organization of the Mushroom Bodies". Philosophical Transactions of the Royal Society of London B. 298 (1091): 309–354. doi:10.1098/rstb.1982.0086.
^ ab López-Riquelme, G.O. (June 2014). "Odotopic afferent representation of the glomerular antennal lobe organization in the mushroom bodies of ants (Hymenoptera: Formicidae): Comparisons between two species". TIP Revista Especializada en Ciencias Químico-Biológicas. 15 (1): 15–31. doi:10.1016/S1405-888X(14)70317-1.
^ Tully, T; Quinn, WG (September 1985). "Classical conditioning and retention in normal and mutant Drosophila melanogaster". J Comp Physiol A. 157 (2): 263–77. doi:10.1007/bf01350033.
^ Gupta, Nitin; Stopfer, Mark (6 October 2014). "A temporal channel for information in sparse sensory coding". Current Biology. 24 (19): 2247–56. doi:10.1016/j.cub.2014.08.021. PMC 4189991. PMID 25264257.
^ Gupta, Nitin; Singh, Swikriti Saran; Stopfer, Mark (2016-12-15). "Oscillatory integration windows in neurons". Nature Communications. 7: 13808. doi:10.1038/ncomms13808. ISSN 2041-1723. PMC 5171764. PMID 27976720.
^ Davis, Ronald (2011). "Traces of Drosophila Memory". Neuron. 70 (1): 8–19. doi:10.1016/j.neuron.2011.03.012.
^ McGuire, Sean; Le, Phuong; Davis, Ronald (August 2001). "The role of Drosophila mushroom body signaling in olfactory memory". Science. 17 (293): 1330–33. doi:10.1126/science.1062622.
^ Bräcker, L. B., Siju, K. P., Varela, N., Aso, Y., Zhang, M., Hein, I., ... & Kadow, I. C. G. (2013). Essential role of the mushroom body in context-dependent CO 2 avoidance in Drosophila. Current Biology, 23(13), 1228-1234.
Chicago
^ Yildizoglu, Tugce; Weislogel, Jan-Marek; Mohammad, Farhan; Chan, Edwin S.-Y.; Assam, Pryseley N.; Claridge-Chang, Adam (2015-12-08). "Estimating Information Processing in a Memory System: The Utility of Meta-analytic Methods for Genetics". PLOS Genet. 11 (12): e1005718. doi:10.1371/journal.pgen.1005718. ISSN 1553-7404. PMC 4672901. PMID 26647168.
^ Akalal, David-Benjamin; Wilson, Curtis; Zong, Lin; Tanaka, Nobuaki; Ito, Kei; Davis, Ronald (September 2006). "Roles for Drosophila mushroom body neurons in olfactory learning and memory". Learning and Memory. 13 (1): 659–68. doi:10.1101/lm.221206. PMC 1783621.
^ Yu, Dinghui; Ponomarev, Artem; Davis, Ronald (May 2004). "Altered representation of the spatial code for odors after olfactory Classical conditioning; memory trace formation by synaptic recruitment". Neuron. 42: 437–49. doi:10.1016/S0896-6273(04)00217-X.
^ Wang, Yalin; Mamira, Akira; Chiang, Ann-shyn; Zhong, Yi (April 2008). "Imaging of an early memory trace in the Drosophila mushroom body". The Journal of Neuroscience. 28 (17): 4368–76. doi:10.1523/jneurosci.2958-07.2008. PMC 3413309. PMID 18434515.
^ Xu, Liu; Davis, Ronald (January 2009). "The GABAergic anterior paired lateral neuron suppresses and is suppressed by olfactory learning" (PDF). Nature Neuroscience. 12 (1): 53–59. doi:10.1038/nn.2235. PMC 2680707. PMID 19043409.
^ Yu, Dinghui; Keene, Alex (December 2005). "Drosophila DPM neurons form a delayed and branch-specific memory trace after olfactory classical conditioning". Cell. 123: 945–57. doi:10.1016/j.cell.2005.09.037. PMID 16325586.
^ Yu, Dinghui; Akalal, Benjamin-David (December 2006). "Drosophila a/b mushroom body neurons form a branch-specific, long-term cellular memory trace after spaced olfactory conditioning". Neuron. 52 (1): 845–55. doi:10.1016/j.neuron.2006.10.030.
^ Akalal, Benjamin-David; Yu, Dinghui (December 2010). "A Late-Phase, Long-Term Memory Trace Forms in the γ Neurons of Drosophila Mushroom Bodies after Olfactory Classical Conditioning". The Journal of Neuroscience. 30 (49): 16699–16708. doi:10.1523/jneurosci.1882-10.2010. PMC 3380342.
^ abcde Tomchik, Seth; Davis, Ronald (November 2009). "Dynamics of Learning-Related cAMP Signaling and stimulus Integration in the Drosophila Olfactory Pathway". Neuron. 64: 510–21. doi:10.1016/j.neuron.2009.09.029. PMC 4080329. PMID 19945393.
^ Abrams, Thomas; Karl, Kevin; Kandel, Eric (September 1991). "Biochemical studies of stimulus convergence during classical conditioning in Aplysia: dual regulation of adenylate cyclase by Ca2+/calmodulin and transmitter". The Journal of Neuroscience. 11 (9): 2655–65.
^ Skoulakis, EM; Kalderon, D; Davis, RL (1993). "Preferential expression in mushroom bodies of the catalytic subunit of protein kinase A and its role in learning and memory". Neuron. 11 (2): 197–201. doi:10.1016/0896-6273(93)90178-t. PMID 8352940.
^ abcdefg Gervasi, Nicolas; Tchènio, Paul; Preat, Thomas (February 2010). "PKA Dynamics in a Drosophila Learning Center: Coincidence Detection by Rutabaga Adenylyl Cyclase and Spatial Regulation by Dunce Phosphodiesterase". Neuron. 65: 516–529. doi:10.1016/j.neuron.2010.01.014. PMID 20188656.
Further reading
.mw-parser-output .refbeginfont-size:90%;margin-bottom:0.5em.mw-parser-output .refbegin-hanging-indents>ullist-style-type:none;margin-left:0.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>ddmargin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none.mw-parser-output .refbegin-100font-size:100%
S.E. Fahrbach (2006). "Structure of the mushroom bodies of the insect brain". Annu. Rev. Entomol. 51: 209–232. doi:10.1006/jtbi.2001.2504. PMID 12183125.
López-Riquelme, Germán Octavio (2008). Hormigas como sistemas modelo para el comportamiento complejo. Bases neurobiológicas de la comunicación química y la división del trabajo en las hormigas (Ph.D.). Universidad Nacional Autónoma de México. doi:10.13140/RG.2.1.3145.1689.
External links
- Standard BeeBrain


