ウイルス
出典は列挙するだけでなく、脚注などを用いてどの記述の情報源であるかを明記してください。記事の信頼性向上にご協力をお願いいたします。(2011年8月) |
| ウイルス | |||
|---|---|---|---|
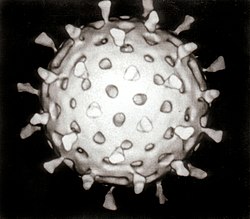 ロタウイルス | |||
分類(ウイルス) | |||
|
ウイルス(ラテン語: virus)は、他生物の細胞を利用して自己を複製させる、極微小な感染性の構造体で、タンパク質の殻とその内部に入っている核酸からなる。生命の最小単位である細胞をもたないので、非生物とされることもある。
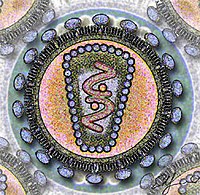
ヒト免疫不全ウイルスの模式図
目次
1 名称
1.1 日本語名
2 特徴
3 歴史
4 一般的な生物との違い
5 構造
5.1 ウイルス核酸
5.2 カプシド
5.3 ヌクレオカプシド
5.4 エンベロープ
6 増殖
6.1 細胞表面への吸着
6.2 細胞内への侵入
6.3 脱殻
6.4 部品の合成
6.5 部品の集合とウイルス粒子の放出
7 宿主に与える影響
7.1 細胞レベルでの影響
7.2 個体レベルでの影響
8 脚注
8.1 注釈
8.2 出典
8.3 参考文献
9 関連項目
10 外部リンク
名称
「ウイルス」は、「毒液」または「粘液」を意味するラテン語 virus に由来して命名された。古代ギリシアのヒポクラテスは病気を引き起こす毒という意味でこの言葉を用いている[要出典]。
日本語名
ヨーロッパの主な言語での発音を以下に列挙する[1]。
| 言語 | 性 | 単数形 | 複数形 | ||
|---|---|---|---|---|---|
| 綴り | 発音 | 綴り | |||
国際音声記号 | カタカナ | ||||
ラテン語 | 中性 | vīrus | [ˈwiːrʊs] | ウィールス | vīra[2][3] |
イタリア語 | 男性 | virus | [ˈvirus] | ヴィールス | virus |
ドイツ語 | 中性 | Virus | [ˈviːrʊs] | ヴィールス | Vira, Viren |
スペイン語 | 男性 | virus | [ˈbiɾus] | ビールス | virus |
フランス語 | 男性 | virus | [viʁys] | ヴィリュス | virus |
英語 | - | virus | [ˈvaɪrəs] | ヴァイラス | viruses (viri[4], vira[5]) |
以上のような発音をもとに、多様な日本語表記が使用された。
| 音写 (語頭の "vi" の音写の違いで分類) | 意訳 | |||||
|---|---|---|---|---|---|---|
| "v"\"vi" | 1音節 | 2音節 | ||||
| 1モーラ | 2モーラ (長音含む) | 3モーラ (長音含む) | 2モーラ | |||
子音 | [b] | ビルス | ビールス | ビイールス | バイラス | 病毒[12][13] 濾過性病原体 |
[v] | ヴィルス | ヴィールス | ヴイールス ヴヰールス | ヴァイラス ヷイラス | ||
[w]([β̞]) | ウィルス | ウィールス | ワイラス | |||
母音 | [u̜]/[ɯ̹] | ウイールス | ウイルス | |||
1953年(昭和28年)に日本ウイルス学会が設立されたのを機に、「ウイルス」という表記が日本語の正式名称として採用された。その後、日本医学会がドイツ語発音に由来する「ビールス」を用いたため混乱があったものの、現在は宿主に関わらず「ウイルス」が正式名称である[14][15]。
なお、ウイルスの細胞外粒子を表す「英: virion」の語には、「ウイリオン」ではなく「ビリオン」の読み表記が定着している。
特徴
ウイルスは細胞を構成単位とせず、自己増殖はできないが、遺伝子を有するという、非生物・生物両方の特性を持っている。現在でも自然科学では生物・生命の定義を行うことができていないため、便宜的に細胞を構成単位とし、代謝、増殖できるものを生物と呼んでいるが、ウイルスは「非細胞性生物」として位置づけられる。あるいは、「生物学的存在」ともいわれる[16]。しかし、遺伝物質を持ち、生物の代謝系を利用して増殖するウイルスは生物と関連があることは明らかである。感染することで宿主の恒常性に影響を及ぼし、病原体としてふるまうことがある。ウイルスを対象として研究する分野はウイルス学と呼ばれる。
遺伝物質の違いから、大きくDNAウイルスとRNAウイルスに分けられる(詳細はウイルスの分類を参照)。真核生物、細菌、古細菌、いずれのドメインにもそれぞれウイルスが発見されており、起源が古いことが示唆されている。細菌に感染するウイルスはバクテリオファージと呼ばれ、分子生物学の初期に遺伝子発現研究のモデル系として多く用いられた。しかし、今日の分子生物学・医学の分野では「ウイルス」という表現は動植物に感染するものを指して用いることが多く、バクテリオファージとは区別して用いることが多い。
歴史
微生物学の歴史は、1674年にオランダのレーウェンフックが顕微鏡観察によって細菌を見出したことに始まり、その後1860年にフランスのルイ・パスツールが生物学や醸造学における意義を、1876年にドイツのロベルト・コッホが医学における意義を明らかにしたことで大きく展開した。特にコッホが発見し提唱した「感染症が病原性細菌によって起きる」という考えが医学に与えた影響は大きく、それ以降、感染症の原因は寄生虫を除いて全て細菌によるものだと考えられていた。
1892年、ロシアのドミトリー・イワノフスキーは、タバコモザイク病の病原が細菌濾過器を通過しても感染性を失わないことを発見、それが細菌よりも微小な、顕微鏡では観察できない存在であることを報告した。またこの研究とは別に、1898年にドイツのフリードリヒ・レフラーとパウル・フロッシュが口蹄疫の病原体の分離を試み、これが同様の存在であることを突き止め、「filterable virus(濾過性病原体)」と呼称した。同年にオランダのマルティヌス・ベイエリンクはイワノフスキーと同様の研究を行って、同じように見出された未知の性質を持つ病原体を「Contagium vivum fluidum(生命を持った感染性の液体)」と呼んだ。
レフラーは濾過性病原体を小さな細菌と考えていたが、ベイエリンクは分子であると考え、これが細胞に感染して増殖すると主張した。彼の主張はすぐには受け入れられなかったが、同様の性質をもった病原体やファージが発見されていくことで、一般にもウイルスの存在が信じられるようになった。その後、物理化学的な性質が徐々に解明され、ウイルスはタンパク質からできていると考えられた。1935年、アメリカのウェンデル・スタンリーがタバコモザイクウイルスの結晶化に成功し、これによってはじめてウイルスは電子顕微鏡によって可視化されることとなった[17]。また彼の発見したこの結晶は、感染能を持っていることを示し、化学物質のように結晶化できる生物の存在は生物学・科学界に衝撃を与えた。彼はこの業績により、1946年にノーベル化学賞を受賞した[18]。スタンリーはウイルスが自己触媒能をもつ巨大なタンパク質であるとしたが、翌年に少量のRNAが含まれることが示された。当時は遺伝子の正体は未解明であり、遺伝子タンパク質説が有力とされていた。当時は、病原体は能動的に病気を引き起こすと考えられていたので、分子ロボット(今で言うナノマシン)のようなもので我々が病気になるということに当時の科学者たちは驚いた。それでも当時はまだ、病原体であるには細菌ほどの複雑な構造、少なくとも自己のタンパク質をコードする遺伝子ぐらいは最低限持っていなくては病原体になりえない、と思われていた。
1952年に行われたハーシーとチェイスの実験は、バクテリオファージにおいてDNAが遺伝子の役割を持つことを明らかにし[19]、これを契機にウイルスの繁殖、ひいてはウイルスの性質そのものの研究が進むようになった。同時に、この実験は生物の遺伝子がDNAであることを示したものと解せられた。
一般的な生物との違い
| 一般的な原核生物 (例:大腸菌) | マイコプラズマ | ナノアルカエウム・エクウィタンス | リケッチア | クラミジア | ファイトプラズマ | ウイルス | |
|---|---|---|---|---|---|---|---|
| 構成単位 | 細胞 | ウイルス粒子 | |||||
| 遺伝情報の担体 | DNA | DNAまたはRNA | |||||
| 増殖様式 | 対数増殖(分裂や出芽) | 一段階増殖 暗黒期の存在 | |||||
ATPの合成 | できる | できない | できる | できない | |||
| タンパク質の合成 | できる | できない | |||||
細胞壁 | ある | ない | ある | ない | |||
| 単独で増殖 | できる | できない (他生物に付着) | できない(偏性細胞内寄生性) | ||||
ウイルスは様々な点で一般的な生物と大きく異なる。
- 非細胞性で細胞質などは持たない。基本的にはタンパク質と核酸からなる粒子である(→ウイルスの構造)。
- 大部分の生物は細胞内部にDNAとRNAの両方の核酸が存在するが、ウイルス粒子内には基本的にどちらか片方だけしかない。
- 他のほとんどの生物の細胞は2nで指数関数的に増殖するのに対し、ウイルスは一段階増殖する。またウイルス粒子が見かけ上消えてしまう「暗黒期」が存在する。
- 代謝系を持たず、単独では増殖できない。他生物の細胞に寄生したときのみ増殖できる。
- 自分自身でエネルギーを産生せず、宿主細胞の作るそれを利用する。
なお4の特徴はウイルスだけに見られるものではなく、リケッチアやクラミジア、ファイトプラズマなど一部の細菌や真核生物にも同様の特徴を示すものがある。
細胞は生きるのに必要なエネルギーを作る製造ラインを持っているが、ウイルスはその代謝を行っておらず、代謝を宿主細胞に完全に依存し、宿主の中でのみ増殖が可能である。彼らに唯一できることは他の生物の遺伝子の中に彼らの遺伝子を入れる事である。厳密には自らを入れる能力も持っておらず、ただ細胞が正常な物質と判別できずウイルスタンパクを増産し病気になる。これらの違いからウイルスは生物学上、生物とは見做されないことも多い。
上記のようにウイルスは生物学上の生物とはされない事が多いが、メガウイルス、ミミウイルスなど、細菌に非常に近い構造を持つウイルスの発見により、少なくとも一部は遺伝子の大部分を捨て去り、寄生に特化した生物の一群由来であろうことが強く示唆されている。一方、レトロウイルスとトランスポゾンの類似性もまた、少なくとも一部のウイルスは機能性核酸が独立・進化したものである可能性を強く示唆している。つまり、「ウイルス」として纏められている物は多元的であり、人為分類群である可能性が非常に高い。
構造

ウイルスの基本構造
(A)エンベロープを持たないウイルス、(B)エンベロープを持つウイルス
ウイルスの基本構造は、粒子の中心にあるウイルス核酸と、それを取り囲むカプシド (capsid) と呼ばれるタンパク質の殻から構成された粒子である。その大きさは小さいものでは数十nmから、大きいものでは数百nmのものまで存在し、他の一般的な生物の細胞(数〜数十µm)の100〜1000分の1程度の大きさである。ウイルス核酸とカプシドを併せたものをヌクレオカプシド (nucleocapsid) と呼ぶ。ウイルスによっては、エンベロープ (envelope) と呼ばれる膜成分など、ヌクレオカプシド以外の物質を含むものがある。これらの構成成分を含めて、そのウイルスにとって必要な構造をすべて備え、宿主に対して感染可能な「完全なウイルス粒子」をビリオンと呼ぶ。
ウイルス核酸
ウイルス核酸は、通常、DNAかRNAのどちらか一方である。すなわち、他の生物が一個の細胞内にDNA(遺伝子として)とRNA(mRNA、rRNA、tRNAなど)の両方の分子を含むのに対して、ウイルスの一粒子にはその片方しか含まれない(ただしDNAと共にRNAを一部含むB型肝炎ウイルスのような例外も稀に存在する)。そのウイルスが持つ核酸の種類によって、ウイルスはDNAウイルスとRNAウイルスに大別される。さらに、それぞれの核酸が一本鎖か二本鎖か、一本鎖のRNAであればmRNAとしての活性を持つか持たないか(プラス鎖RNAかマイナス鎖RNAか)、環状か線状か、などによって細かく分類される。ウイルスのゲノムは他の生物と比べてはるかにサイズが小さく、またコードしている遺伝子の数も極めて少ない。例えば、ヒトの遺伝子が数万あるのに対して、ウイルスでは3〜100個ほどだと言われる。
ウイルスは基本的にタンパク質と核酸からなる粒子であるため、ウイルスの複製(増殖)のためには少なくとも
- タンパク質の合成
- ウイルス核酸の複製
- 1. 2.を行うために必要な、材料の調達とエネルギーの産生
が必要である。しかしほとんどのウイルスは、1や3を行うのに必要な酵素の遺伝情報を持たず、宿主細胞の持つタンパク合成機構や代謝、エネルギーを利用して、自分自身の複製を行う。ウイルス遺伝子には自分の遺伝子(しばしば宿主と大きく異なる)を複製するための酵素の他、宿主細胞に吸着・侵入したり、あるいは宿主の持つ免疫機構から逃れるための酵素などがコードされている。
ウイルスによっては、カプシドの内側に、核酸と一緒にカプシドタンパク質とは異なるタンパク質を含むものがある。このタンパク質とウイルス核酸を合わせたものをコアと呼び、このタンパク質をコアタンパク質と呼ぶ。
カプシド
カプシド (capsid) は、ウイルス核酸を覆っているタンパク質であり、ウイルス粒子が細胞の外にあるときに内部の核酸をさまざまな障害から守る「殻」の役割をしていると考えられている。ウイルスが宿主細胞に侵入した後、カプシドが壊れて(脱殻、だっかく)内部のウイルス核酸が放出され、ウイルスの複製がはじまる。
カプシドは、同じ構造を持つ小さなタンパク質(カプソマー)が多数組み合わさって構成されている。この方式は、ウイルスの限られた遺伝情報量を有効に活用するために役立っていると考えられている。小さなタンパク質はそれを作るのに必要とする遺伝子配列の長さが短くてすむため、大きなタンパク質を少数組み合わせて作るよりも、このように小さいタンパク質を多数組み合わせる方が効率がよいと考えられている。
ヌクレオカプシド

ヌクレオカプシドの対称性(左)正二十面体様(中)らせん構造(右)構造の複雑なファージ
ウイルス核酸とカプシドを合わせたものをヌクレオカプシド (nucleocapsid) と呼ぶ。エンベロープを持たないウイルスではヌクレオカプシドはビリオンと同じものを指す。言い換えればヌクレオカプシドは全てのウイルスに共通に見られる最大公約数的な要素である。
ヌクレオカプシドの形はウイルスごとに決まっているが、多くの場合、正二十面体様の構造、またはらせん構造をとっており、立体対称性を持つ。ただし、天然痘の原因であるポックスウイルスやバクテリオファージなどでは、ヌクレオカプシドは極めて複雑な構造であり、単純な対称性は持たない。
エンベロープ
詳細は「エンベロープ (ウイルス)」を参照
エンベロープ (envelope) は、単純ヘルペスウイルスやインフルエンザウイルス、ヒト免疫不全ウイルスなど一部のウイルス粒子に見られる膜状の構造のこと。これらのウイルスにおいて、エンベロープはウイルス粒子(ビリオン)の最も外側に位置しており、ウイルスの基本構造となるウイルスゲノムおよびカプシドタンパク質を覆っている。エンベロープの有無はウイルスの種類によって決まっており、分離されたウイルスがどの種類のものであるかを鑑別する際の指標の一つである。
エンベロープは、ウイルスが感染した細胞内で増殖し、そこから細胞外に出る際に細胞膜あるいは核膜などの生体膜を被ったまま出芽することによって獲得されるものである。このため、基本的には宿主細胞の脂質二重膜に由来するものであるが、この他にウイルス遺伝子にコードされている膜タンパク質の一部を細胞膜などに発現した後で膜と一緒にウイルス粒子に取り込み、エンベロープタンパク質としてビリオン表面に発現させている。これらのエンベロープタンパク質には、そのウイルスが宿主細胞に吸着・侵入する際に細胞側が持つレセプターに結合したり、免疫などの生体防御機能を回避したりなど、さまざまな機能を持つものが知られており、ウイルスの感染に重要な役割を果たしている。
細胞膜に由来するエンベロープがあるウイルスでは、エンベロープタンパク質が細胞側のレセプターに結合した後、ウイルスのエンベロープと細胞膜とが膜融合を起こすことで、エンベロープ内部に包まれていたウイルスの遺伝子やタンパク質を細胞内に送り込む仕組みのものが多い。
エンベロープはその大部分が脂質から成るためエタノールや有機溶媒、石けんなどで処理すると容易に破壊することができる。このため一般にエンベロープを持つウイルスは、消毒用アルコールでの不活化が、エンベロープを持たないウイルスに比べると容易である。
増殖

細胞(左)とウイルス(右)の増殖様式
ウイルスは、それ自身単独では増殖できず、他の生物の細胞内に感染して初めて増殖可能となる。このような性質を偏性細胞内寄生性と呼ぶ。また、一般的な生物の細胞が2分裂によって 2n で対数的に数を増やす(対数増殖)のに対し、ウイルスは1つの粒子が、感染した宿主細胞内で一気に数を増やして放出(一段階増殖)する。また感染したウイルスは細胞内で一度分解されるため、見かけ上ウイルス粒子の存在しない期間(暗黒期)がある。
ウイルスの増殖は以下のようなステップで行われる。
細胞表面への吸着 → 細胞内への侵入 → 脱殻(だっかく) → 部品の合成 → 部品の集合 → 感染細胞からの放出
細胞表面への吸着
ウイルス感染の最初のステップはその細胞表面に吸着することである。ウイルスが宿主細胞に接触すると、ウイルスの表面にあるタンパク質が宿主細胞の表面に露出しているいずれかの分子を標的にして吸着する。このときの細胞側にある標的分子をそのウイルスに対するレセプターと呼ぶ。ウイルスが感染するかどうかは、そのウイルスに対するレセプターを細胞が持っているかどうかに依存する。代表的なウイルスレセプターとしては、インフルエンザウイルスに対する気道上皮細胞のシアル酸糖鎖や、ヒト免疫不全ウイルスに対するヘルパーT細胞表面のCD4分子などが知られている。
細胞内への侵入
細胞表面に吸着したウイルス粒子は、次に実際の増殖の場になる細胞内部へ侵入する。侵入のメカニズムはウイルスによってさまざまだが、代表的なものに以下のようなものがある。
エンドサイトーシスによる取り込み- 細胞自身が持っているエンドサイトーシスの機構によって、エンドソーム小胞として細胞内に取り込まれ、その後でそこから細胞質へと抜け出すもの。エンベロープを持たないウイルスの多くや、インフルエンザウイルスなどに見られる。
- 膜融合
- 吸着したウイルスのエンベロープが細胞の細胞膜と融合し、粒子内部のヌクレオカプシドが細胞質内に送り込まれるもの。多くの、エンベロープを持つウイルスに見られる。
- 能動的な遺伝子の注入
- Tファージなどのバクテリオファージに見られ、吸着したウイルスの粒子から尾部の管を通してウイルス核酸が細胞質に注入される。注入とは言っても、ウイルス粒子の尾部が細菌の細胞壁を貫通した後の遺伝子の移動は、細菌細胞が生きていないと起こらないため、細菌の細胞自体の作用によって吸い込まれるのではないかと言われている。
脱殻
細胞内に侵入したウイルスは、そこで一旦カプシドが分解されて、その内部からウイルス核酸が遊離する。この過程を脱殻と呼ぶ。脱殻が起こってから粒子が再構成されるまでの期間は、ビリオン(感染性のある完全なウイルス粒子)がどこにも存在しないことになり、この時期を暗黒期、あるいは日食や月食になぞらえてエクリプス期(eclipse period)と呼ぶ。
部品の合成
脱殻により遊離したウイルス核酸は、次代のウイルス(娘ウイルス)の作成のために大量に複製されると同時に、さらにそこからmRNAを経て、カプソマーなどのウイルス独自のタンパク質が大量に合成される。すなわちウイルスの合成は、その部品となる核酸とタンパク質を別々に大量生産し、その後で組み立てるという方式で行われる。
ウイルス核酸は宿主細胞の核酸とは性質的に異なる点が多いために、その複製は宿主の持つ酵素だけではまかなえないため、それぞれのウイルスが独自に持つDNAポリメラーゼ、RNAポリメラーゼなど、転写・複製に関わる酵素が使われる。また逆転写酵素を持つレトロウイルスでは、宿主のDNAに自分の遺伝子を組み込むことで、宿主のDNA複製機構も利用する。
タンパク質の合成には、そのタンパク質をコードするmRNAを作成するためにウイルス独自の酵素を必要とする場合がある。mRNAからタンパク質への翻訳は、宿主細胞の持つ、リボソームなどのタンパク質合成系を利用して行われる。
部品の集合とウイルス粒子の放出
別々に大量生産されたウイルス核酸とタンパク質は細胞内で集合する。最終的にはカプソマーがウイルス核酸を包み込み、ヌクレオカプシドが形成される。この機構はウイルスによってまちまちであり、まだ研究の進んでないものも多い。細胞内で集合したウイルスは、細胞から出芽したり、あるいは感染細胞が死ぬことによって放出される。このときエンベロープを持つウイルスの一部は、出芽する際に被っていた宿主の細胞膜の一部をエンベロープとして獲得する。
宿主に与える影響
ウイルスによる感染は、宿主となった生物に細胞レベルや個体レベルでさまざまな影響を与える。その多くの場合、ウイルスが病原体として作用し、宿主にダメージを与えるが、一部のファージやレトロウイルスなどに見られるように、ウイルスが外来遺伝子の運び屋として作用し、宿主の生存に有利に働く例も知られている。
細胞レベルでの影響
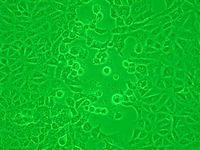
細胞変性効果(円形化)培養フラスコの底に敷石状に生育している培養細胞がウイルスの感染によって円く変形し、やがてフラスコからはがれてプラーク(空隙、写真中央)を形成する。

細胞変性効果(合胞体)敷石状に生育した培養細胞同士がウイルス感染によって細胞膜の融合を起こし、細胞核が中央に凝集して(写真中央)多核巨細胞様の形態になる。
ウイルスが感染して増殖すると、宿主細胞が本来自分自身のために産生・利用していたエネルギーや、アミノ酸などの栄養源がウイルスの粒子複製のために奪われ、いわば「ウイルスに乗っ取られた」状態になる。
これに対して宿主細胞はタンパク質や遺伝子の合成を全体的に抑制することで抵抗しようとし、一方でウイルスは自分の複製をより効率的に行うために、さまざまなウイルス遺伝子産物を利用して、宿主細胞の生理機能を制御しようとする。またウイルスが自分自身のタンパク質を一時に大量合成することは細胞にとって生理的なストレスになり、また完成した粒子を放出するときには宿主の細胞膜や細胞壁を破壊する場合もある。このような原因から、ウイルスが感染した細胞ではさまざまな生理的・形態的な変化が現れる。
この現象のうち特に形態的な変化を示すものを細胞変性効果 (cytopathic effect, CPE) と呼ぶ。ウイルスによっては、特定の宿主細胞に形態的に特徴のある細胞変性効果を起こすものがあり、これがウイルスを鑑別する上での重要な手がかりの一つになっている。代表的な細胞変性効果としては、細胞の円形化・細胞同士の融合による合胞体 (synsitium) の形成・封入体の形成などが知られる。
さまざまな生理機能の変化によって、ウイルスが感染した細胞は最終的に以下のいずれかの運命を辿る。
- ウイルス感染による細胞死
- ウイルスが細胞内で大量に増殖すると、細胞本来の生理機能が破綻したり細胞膜や細胞壁の破壊が起きる結果として、多くの場合、宿主細胞は死を迎える。ファージ感染による溶菌現象もこれにあたる。多細胞生物の細胞では、ウイルス感染時に細胞周期を停止させたり、MHCクラスIなどの抗原提示分子を介して細胞傷害性T細胞を活性化して、アポトーシスを起こすことも知られている。感染した細胞が自ら死ぬことで周囲の細胞にウイルスが広まることを防いでいると考えられている。
- 持続感染
- ウイルスによっては、短期間で大量のウイルスを作って直ちに宿主を殺すのではなく、むしろ宿主へのダメージが少なくなるよう少量のウイルスを長期間に亘って持続的に産生(持続感染)するものがある。宿主細胞が増殖する速さと、ウイルス複製による細胞死の速さが釣り合うと持続感染が成立する。テンペレートファージによる溶原化もこれにあたる。持続感染の中でも、特にウイルス複製が遅くて、ほとんど粒子の複製が起こっていない状態を潜伏感染と呼ぶ。
- 細胞の不死化とがん化
- 多細胞生物に感染するウイルスの一部には、感染した細胞を不死化したり、がん化したりするものが存在する。このようなウイルスを腫瘍ウイルスあるいはがんウイルスと呼ぶ。ウイルスが宿主細胞を不死化あるいはがん化させるメカニズムはまちまちであるが、宿主細胞が感染に抵抗して起こす細胞周期停止やアポトーシスに対抗して、細胞周期を進行させたりアポトーシスを抑制する遺伝子産物を作る場合(DNAがんウイルス)や、細胞の増殖を活性化する場合、またレトロウイルスでは宿主のゲノムにウイルス遺伝子が組み込まれる際、がん抑制遺伝子が潰された結果、がん化することも知られている。
個体レベルでの影響
ウイルス感染は、細胞レベルだけでなく多細胞生物の個体レベルでも、さまざまな病気を引き起こす。このような病気を総称してウイルス感染症と呼ぶ。インフルエンザや天然痘、麻疹、風疹、後天性免疫不全症候群(AIDS)などの病気がウイルス感染症に属しており、これらのウイルスはしばしばパンデミックを引き起こして人類に多くの犠牲者を出した。
また、動物ではウイルス感染が起きると、それに抵抗して免疫応答が引き起こされる。血液中や粘液中のウイルス粒子そのものに対しては、ウイルスに対する中和抗体が作用する(液性免疫)ことで感染を防ぐ。感染した後の細胞内のウイルスに対しては抗体は無効であるが、細胞傷害性T細胞やNK細胞などが感染細胞を殺す(細胞性免疫)ことで感染の拡大を防ぐ。免疫応答はまた、特定のウイルス感染に対して人工的に免疫を付与するワクチンによっても産生され得る。AIDSやウイルス性肝炎の原因となるものを含む一部のウイルスは、これらの免疫応答を回避し、慢性感染症を引き起こす。
ウイルス感染症における症状の中には、ウイルス感染自体による身体の異常もあるが、むしろ発熱、感染細胞のアポトーシスなどによる組織傷害のように、上記のような免疫応答を含む、対ウイルス性の身体の防御機構の発現自体が健康な身体の生理機構を変化させ、さらには身体恒常性に対するダメージともなり、疾患の症状として現れるものが多い。
脚注
注釈
出典
^ Definition of "virus" > Translations for 'virus' (Collins English Dictionary)
^ William T. Stearn: Botanical Latin. History, Grammar, Syntax, Terminology and Vocabulary. Third edition, David & Charles, 1983. 「Virus: virus (s.n. II), gen. sing. viri, nom. pl. vira, gen. pl. vīrorum (to be distinguished from virorum, of men).」
^ Pons: virus
^ e.g. Michael Worboys: Cambridge History of Medicine: Spreading Germs: Disease Theories and Medical Practice in Britain, 1865-1900, Cambridge University Press, 2000, p. 204
^ e.g. Karsten Buschard & Rikke Thon: Diabetic Animal Models. In: Handbook of Laboratory Animal Science. Second Edition. Volume II: Animal Models, edited by Jann Hau & Gerald L. Van Hoosier Jr., CRC Press, 2003, p. 163 & 166
^ 文章表現辞典(東京堂出版 1965年) p.48
^ 學術集談會演説要旨 (PDF) (1937年3月25日發表)
^ 植物のヷイラス病(信州大学 学術情報オンラインシステムSOAR)
^ インフルエンザ・ヴイールスの赤血球による收着(大阪大学)
^ 日本脳炎ウイールスの赤血球凝集反応(第1報)(千葉大学)
^ 家兎ニヨル發疹チフス病毒ノ實驗的研究(北里研究所)
^ 日本細菌学会が意訳。中国語に取り入れられ、現在でも使用されている。
^ 労働安全衛生規則(昭和47年9月30日労働省令第32号)第61条第1項第1号に「病毒伝ぱのおそれのある伝染性の疾病にかかった者」とあるが、この場合の伝染性疾患とは結核を指すとされている(労働安全衛生規則等の一部を改正する省令の施行について 平成12年3月30日・基発第207号より)。すなわち、この場合の病毒はウイルスではなく、細菌である結核菌を指す。
^ http://jsv.umin.jp/about_jsv/about_jsv.html
^ 日本植物病理学会編 (1995). 植物病理学事典. 養賢堂. pp. p.91.
^ マシューズ、ホルダ、アハーン『カラー生化学』西村書店刊、2003年5月15日発行(16ページ)
^ 「人類と感染症の歴史 未知なる恐怖を越えて」p4 加藤茂孝 丸善出版 平成25年3月30日発行
^ http://www.nobelprize.org/nobel_prizes/chemistry/laureates/1946/stanley-facts.html 「Wendell M. Stanley - Facts」ノーベル賞財団 2015年10月4日閲覧
^ http://www.sci.toho-u.ac.jp/biomol/glossary/bio/Hershey_Chase_experiment.html 「ハーシーとチェイスの実験(Hershey-Chase experiment)」東邦大学理学部生物分子科学科 2015年10月4日閲覧
参考文献
- 『STEP内科 2 感染症・血液』松岡健 他 監修 海馬書房 1998年11月27日 初版 P.107〜P.109
- 『医科ウイルス学』大里外誉郎編集、南江堂 1992年12月
- 『医学ウイルス学』David O. White, Frank J. Fenner
- 『ウイルス図鑑』保坂康弘ほか
- 『医学細菌学』中野昌康 菜根出版
関連項目
- バクテリオファージ
- ウイロイド
- サテライトウイルス
- ウイルスフリー
- ウイルス学
- ウイルス学の歴史
- ウイルスの社会史
- ウイルス療法
- ウイルス進化論
- 生物兵器
- 病気
後天性免疫不全症候群 (AIDS)
重症急性呼吸器症候群(新型肺炎、SARS)- インフルエンザ
- ウイルス性肝炎
- 生物以外
- コンピュータウイルス
- ミーム#マインド・ウイルス
外部リンク
- 日本ウイルス学会
- ウイルス図鑑
- 世界大百科事典 第2版『ウイルス』 - コトバンク
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||


