モササウルス科
| モササウルス科 Mosasauridae | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
生息年代: 白亜紀後期 PreЄ Є O S D C P T J K Pg N | |||||||||||||||||||||||||||||||||
 マーストリヒト自然史博物館に展示されているモササウルスの骨格 | |||||||||||||||||||||||||||||||||
地質時代 | |||||||||||||||||||||||||||||||||
白亜紀 | |||||||||||||||||||||||||||||||||
分類 | |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
学名 | |||||||||||||||||||||||||||||||||
Mosasauridae Gervais, 1853 | |||||||||||||||||||||||||||||||||
亜科 | |||||||||||||||||||||||||||||||||
本文参照 |
モササウルス科とは白亜紀後期に繁栄した有鱗目トカゲ亜目オオトカゲ上科に属する、海生のトカゲからなるグループである。まれに海トカゲ類と呼ばれることもある。本項目では模式属であるモササウルスを含めた科全体の概要について解説する。
モササウルスの化石は1764年にオランダのマーストリヒトの石灰岩採石場で発見された。その後のジョルジュ・キュヴィエらの研究によって、この化石が絶滅した大型の海生爬虫類のものであることが判明した。
現在の分岐分類学では顎の構造などから、モササウルス科はトカゲ亜目の中でもヘビに最も近縁なグループであると考えられており[1]、有鱗目の種分化を考えるうえで非常に注目されている分類群である。
目次
1 概要
2 軟組織
3 生息環境
4 研究史
5 分類と系統
5.1 系統
6 モササウルス科の祖先
7 関連項目
8 参考文献
9 外部リンク
概要

チャールズ・ナイトによる背びれが描かれたティロサウルスの復元。気管軟骨が背鰭を支えていた軟骨であると誤認され、この復元図が描かれた[2][3]。

尾びれが三日月型のプラテカルプスの復元。
モササウルス科は白亜紀中頃の魚竜、プリオサウルス類、海生ワニ類の絶滅(南太平洋で起きた巨大海底火山活動による海洋無酸素事変等の海洋環境の悪化が原因とされる。この火山活動によって現在のニューギニア北方沖にあるオントンジャワ海台が形成された)に乗じて空白になった生態域に進出し、白亜紀後期のチューロニアンからマーストリヒチアンにかけての2000万年間、海中の頂点捕食者として繁栄を遂げた。温暖な大陸棚の浅海での生活によく適応し、汎世界的に分布を広げた。特筆されるのは、モササウルス類は胎生であり卵を陸上に産む必要がなく、ウミガメよりも海中生活に適応していたことである。
モササウルス科最小の種は体長3-3.5mのカリノデンスである。この種は球根状の頑丈な歯を持ち、海岸線近くの浅い海底で貝や棘皮動物を食べていた。ほとんどのモササウルス類はこれよりも大きく、ティロサウルスは最大体長17.5m、上述のマーストリヒトから発見されたモササウルスの最初の標本も体長18m前後に達するとみられる。大型モササウルス類の体長はほかの有鱗目を大きく引き離しており、彼らは史上最大級の有鱗目であるといえる(参考までにティタノボアの全長:12-15m、アミメニシキヘビの全長:9.9m、ハナブトオオトカゲの全長:4.75m)。
モササウルスの体型は現生のオオトカゲに似ているが、遊泳生活に適応するためにより胴長で流線型になった。肋骨は短くなり、指骨が伸びて手足は櫂のような形をしている。尾は垂直方向に幅広くなり、これで推進力を生み出した。尾を使った泳ぎ方はウナギやウミヘビなどと共通する。しかし最新の研究では、モササウルスの尾には、サメや魚竜のように大きな三日月型の鰭があったという説が唱えられている(左上図参照)。モササウルスの筋肉をみると、ヘビのような体全体をくねらせる泳ぎ方よりもむしろ、尾鰭で力強く水を蹴って推進力を生み出す泳ぎ方のほうが、水理学的にかなう泳ぎ方である[4]。モササウルスは待ち伏せ型の狩りを行い、尾鰭の強力な推進力で獲物を急襲していた可能性もある。
モササウルスは二重関節の顎ととても柔軟な頭蓋骨を持っていた。この形質は、彼らが大型の獲物をいっきに丸呑みにするための適応であると考えられる。このヘビのような貪欲な捕食方法は、モササウルスの体内からほとんど噛み砕かれていない獲物の化石が見つかっていることからも裏付けられている。また、モササウルス類は海中生物を手当り次第に食べていたと考えられる。サウスダコタから発見されたティロサウルスの体内からは胃内容物として、ヘスペロルニス(ウに似た海鳥)、硬骨魚類、サメ、より小型のモササウルス類(クリダステス)の化石が見つかった。ちなみに、モササウルス類の骨は大量のサメの歯の間から見つかった。
モササウルスのさまざまな特徴(二列に並ぶ口蓋歯、緩く連結した二重関節の顎、短い肋骨、体をくねらせる遊泳・移動方法など)から、多くの研究者はモササウルスがヘビと共通の祖先から分化したと考えている。この学説は1869年にエドワード・ドリンカー・コープによってはじめて提唱され、彼はヘビとモササウルスを”Pythonomorpha”という分類群に統一しようとした。この学説はながく忘れ去られていたが、1990年代に入って再び脚光を浴びた[5][6]。
軟組織

ティロサウルスの鱗の印象化石(KUVP-1075)
かなりの数のモササウルス科の化石が世界中から産出しており、表皮に関する情報は比較的早くから知られていた。世界中から見つかったモササウルス類の化石のうち、いくつかが鱗の印象化石を残していた。もしかするとすでに鱗の印象が残るモササウルスの化石は見つかっていたのかもしれないが、このようなデリケートな部位が化石に残るとは考えられておらず、長い間見過ごされてきたのかもしれない。
モササウルス類の外皮の特徴は長い間、アメリカのカンザス州コーヴ郡の上部サントニアン〜下部カンパニアンから見つかったティロサウルスの骨格標本(KUVP-1075)にもとづいて研究がなされてきた(右図)[7] 。ヨルダンのハラナにある Muwaqqar層から保存状態のよいモササウルス類の化石が見つかった。この化石は、手足の指骨の間の薄い皮膜の部分を含めて、ヘビのようにオーバーラップしたダイヤ型の細かい鱗で覆われていた[8]。現生の爬虫類のようにモササウルス類も、体の場所によって鱗の大きさや形が異なっていたことが分かっている。ハラナの標本からは2種類の鱗が識別された[8]。 稜線がある鱗は体の上部を覆い、表面が滑らかな鱗は体の下部を覆っていた。奇襲型の捕食者は待ち伏せ型の狩りを行うことから、モササウルス類は光を反射しにくい稜線がある鱗を利用していたと考えられる[9]。

プラテカルプス(LACM 128319)の頭部と頸部から見つかった軟組織。 気管軟骨が写真の下部に写っている。
最近、皮膚組織だけではなく内部組織まで保存された、きわめて状態のよいプラテカルプスの標本が見つかった。体内には心臓、肺、腎臓とおぼしき赤い組織が残されていた。さらに気管軟骨と網膜とおぼしき組織までが保存されていた。“腎臓”は腹部のはるか前方にあり、この配置はオオトカゲよりもむしろクジラ類に似ている。オオトカゲもふくめた現生の爬虫類では、気管支は途中で二股に分岐して肺に繋がるが、モササウルス類ではクジラのように気管支は左右独立して肺まで平行に配列する。これらの特徴はモササウルスが水中生活へ移行したことによる、内部形態の変化であると考えられる[4]。
さらに2011年には、モササウルス科のプログナトドンの化石から白亜紀のものとされるコラーゲンが見つかった[10]。
生息環境
IRSNB 1624から見つかった繊維状の組織
白亜紀の海水準は現在よりもはるかに高く、世界各地で海進を引き起こした。モササウルス科の化石はオランダ、デンマーク、ポルトガル、スウェーデン、イギリス[11]、アンゴラ、モロッコ、ニュージーランド、メキシコ、ペルー、はては南極沖のヴェガ島からも見つかっている。なお、従来ニュージーランド産の”恐竜化石”とされたものの多くが実はモササウルス類、ないし首長竜の化石であったことが判明している。北アメリカでは、かつてのニオブララ海の分布域である、カナダのマニトバ州、サスカチュワン州[12]、アメリカ中西部を中心にモササウルス類がひろく見つかっている[13][14]。ちなみに、日本からは北海道三笠市からモササウルス科タニファサウルス属の一種、エゾミカサリュウが見つかっている。
長年、海にのみ住んでいると考えられていたが、2012年12月19日付のオンライン科学雑誌プロスワンに、ハンガリー自然史博物館のチームが、かつて川だった8400万年前の地層からモササウルス類の化石を発見したと発表した。淡水で生活する種も居た可能性が考えられている[15]。
研究史

1770〜1774年にオランダ・マーストリヒトで見つかったモササウルスの頭蓋骨
最初に報告されたモササウルス類の化石は、1764年にオランダのマーストリヒト市近郊の採石場の鉱夫達によって見つかったモササウルスの部分的な頭蓋骨である。モササウルス化石の発見は恐竜化石の発見(イグアノドン:1822年、メガロサウルス:1824年)よりも早いが、有名な恐竜の陰に隠れてあまり知られていない。しかし啓蒙時代に入り、知識人たちの自然科学への興味が高まるにつれて、ヨーロッパ各地で見つかっていた大型化石はじつは太古の絶滅生物の遺骸なのではないかという関心が持たれるようになった。そのような中で、モササウルスの第二の標本が発見される。
1770年から1774年の間に、軍医で化石コレクターであったJohann Leonard Hoffmannはモササウルスの第二標本を当時の一流の科学者達に引き合わせ、この化石の存在が有名になった。当時のこの化石を管理していたのはマーストリヒト大聖堂の司祭、Goddingであった。
1794年に、マーストリヒトがフランス軍によって占領された時(フランス革命戦争)、モササウルスの第二標本は戦利品として600本のワインボトルとともにパリに持ち去られた。
当初、その化石は魚、ワニ、あるいはマッコウクジラであるとさまざまに解釈されていたが、1799年になってオランダ人科学者Adriaan Gilles Camperがはじめてこの化石とトカゲの共通点を指摘した。1808年、ジョルジュ・キュヴィエは比較解剖学の見地からこの結論を支持し、モササウルスが絶滅した未知の海生爬虫類であることを見抜いた。この研究は古脊椎動物に対する本格的な比較解剖学的研究の最初の例である。彼は1822年にそれまでle Grand Animal fossile de Maëstricht(マーストリヒトの大型動物の化石)と呼ばれていたこの化石をMosasaurus(マース川のトカゲの意)と命名した。この経験はのちの彼の学説(天変地異説)に大きな影響を与えた。
その後、1829年にはこの化石に完全な形での学名Mosasaurus hoffmanni(Hoffmann氏のマース川のトカゲの意)を与えた。他にモササウルスの化石の断片とおぼしきものは以前からマーストリヒトで見つかってはいたが、19世紀までに学術的な意義は認識されなかった。ハールレムのテイラーズ博物館には1790年から展示されていた。マーストリヒトの石灰岩層はモササウルスの発見で一躍有名になった。そして、白亜紀の最後の時代はマーストリヒチアンと呼ばれるようになった。
分類と系統
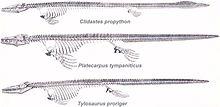
Williston (1898)より、カンザス州から多く産出するモササウルス類3種;クリダステス、プラテカルプス、ティロサウルス

タニファサウルス

クリダステス

プログナトドン
- アイギアロサウルス科
アイギアロサウルス Aigialosaurus
- モササウルス亜科
ダラサウルス Dallasaurus
クリダステス Clidastes
モアナサウルス Moanasaurus
アンフェケプビス Amphekepubis
プロトサウルス Plotosaurus
モササウルス Mosasaurus
グロビデンス Globidens
プログナトドン Prognathodon
プレシオティロサウルス Plesiotylosaurus
カリノデンス Carinodens
- ティロサウルス亜科
ハイノサウルス Hainosaurus
ティロサウルス Tylosaurus
タニファサウルス Taniwhasaurus - エゾミカサリュウを含む
- プリオプラテカルプス亜科
エクテノサウルス Ectenosaurus
プレシオプラテカルプス Plesioplatecarpus
アンゴラサウルス Angolasaurus
プラテカルプス Platecarpus
プリオプラテカルプス Plioplatecarpus
セルマサウルス Selmasaurus
イグダマノサウルス Igdamanosaurus
- ハリサウルス亜科
エオナタトル Eonatator
ハリサウルス Halisaurus
- テティサウルス亜科
パンノニアサウルス Pannoniasaurus
テティサウルス Tethysaurus
ヤグアラサウルス Yaguarasaurus
ルッセロサウルス Russellosaurus
- Incertae sedis
リオドン Liodon
ゴロニオサウルス Goronyosaurus
プルリデンス Pluridens
コウリソドン Kourisodon
系統
モササウルス科と近縁の分類群を含むクラドグラム。Aaron et al.(2012)等による[16][15]。
| モササウルス上科 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
モササウルス科の祖先

モササウルス科の祖先形と目されるアイギアロサウルス
上述したモササウルス科とヘビの共通した特徴から、両者は共通の海生の祖先を持っていると考えられている。白亜紀前期のヨーロッパに生息していたアイギアロサウルスは現生のオオトカゲに似た体型の半水生のトカゲであり、モササウルス科はこのような生物から進化したのかもしれない。2005年に発見されたダラサウルスはモササウルス科の中でも原始的な形状を残しており、陸生のオオトカゲと海生のモササウルス科の中間的な存在であるとされる[17]。
一方、2006年には南アメリカから白亜紀後期の地中性の原始的なヘビである”ナジャシュ”が発見されたことによって、モササウルス=ヘビの海中起源説に対する反論もなされている。
関連項目
ニオブララ層:アメリカ中西部に分布する白亜系の海成層。モササウルス類の化石を多産する。- オオトカゲ上科
- ヘビ亜目
- 有鱗目
参考文献
^ Lee MSY (1997-01-29), “The phylogeny of varanoid lizards and the affinities of snakes”, Philos Trans R Soc Lond B Biol Sci. 352 (1349): 53–91, doi:10.1098/rstb.1997.0005, PMC 1691912, http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1691912
^ http://www.oceansofkansas.com/Osborn1899.html
^ http://www.oceansofkansas.com/Williston98.html- ^ abLindgren, J.; Caldwell, M.W.; Konishi, T.; and Chiappe, L.M. (2010), Farke, Andrew Allen, ed., “Convergent evolution in aquatic tetrapods: Insights from an exceptional fossil mosasaur”, PLoS ONE 5 (8): e11998, doi:10.1371/journal.pone.0011998, PMC 2918493, PMID 20711249, http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2918493
^ Palaeos Vertebrates 260.100 Pythonomorpha: Pythonomorpha
^ Mosasaurs: Last of the Great Marine Reptiles
^ Snow, F. H. (1878), “On the dermal covering of a mosasauroid reptile”, Transactions of the Kansas Academy of Science 6: 54–58- ^ abKaddumi, H.F. (2009), “On the latest scale coverings of mosasaurs (Squamata: Mosasauridae) from the Harrana Fauna in addition to the description of s new species of Mosasaurus”, Fossils of the Harrana Fauna and the Adjacent Areas, Amman: Eternal River Museum of Natural History, pp. 80–94
^ Massare, J. A. (1987), “Tooth morphology and prey preference of Mesozoic marine reptiles”, Journal of Vertebrate Paleontology 7 (2): 121–137, doi:10.1080/02724634.1987.10011647
^ http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0019445
^ ST. JAMES' PIT DISTRICT (PDF)
^ http://www.discoverfossils.com
^ Michael J. Everhart (2005), “Chapter 9: Enter the Mosasaurs”, Oceans of Kansas: a natural history of the western interior sea, Bloomington: Indiana University Press, ISBN 0-253-34547-2
^ Getman, Myron RC (1994), Occurrences of Mosasaur and other reptilian fossil remains from the Fox Hills Formation (Maastrichtian: late Cretaceous) of North Dakota, St. Lawrence University Dept. of Geology theses- ^ abLászló Makádi, Michael W. Caldwell, Attila Ősi (2012). “The First Freshwater Mosasauroid (Upper Cretaceous, Hungary) and a New Clade of Basal Mosasauroids”. PLoS ONE 7 (12): e51781. doi:10.1371/journal.pone.0051781.
^ Aaron R. H. Leblanc, Michael W. Caldwell and Nathalie Bardet (2012). “A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics”. Journal of Vertebrate Paleontology 32 (1): 82–104. doi:10.1080/02724634.2012.624145.
^ Dallasaurus / Ancient Mosasaur - Dallasaurus - SMU
外部リンク
- Palaeos: Vertebrates: Mosasaurs
- BBC Science and Nature: Mosasaurs
Mike Everhart and David Lewis, "Mesozoic marine monsters of the Mangahouanga": New Zealand fossil fauna
Mike Everhart, "A day in the life of a Mosasaur": life in the Sea of Kansas, illus. by Carl Buell
Mike Everhart, "Mosasaurus hoffmani" until 1829.
Mosasaurus maximus mounted skeleton at University of Texas Memorial Museum- Canadian Fossil Discovery Centre
"The Mosasaur of Maastricht" by Hennie Reuvers in Crossroads web magazine- "Mosasaurs terrorized Cretaceous rivers" Planet Earth online
- Georgia Southern University Museum Mosasaur Exhibit


