カブトガニ綱
このページの名前に関して「節口綱」もしくは「腿口類」への改名が提案されています。 議論はこのページのノートを参照してください。(2018年11月) |
| カブトガニ綱 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
生息年代: 467.3–0 Ma PreЄ Є O S D C P T J K Pg N オルドビス紀(ダリウィル期)– 現世[1][2] | ||||||||||||
  エルンスト・ヘッケルによるミナミカブトガニ(上)とウミサソリ(下)のイラスト | ||||||||||||
地質時代 | ||||||||||||
オルドビス紀(ダリウィル期) - 現世[1] | ||||||||||||
分類 | ||||||||||||
| ||||||||||||
学名 | ||||||||||||
Merostomata Woodward, 1866 | ||||||||||||
目 | ||||||||||||
|
カブトガニ綱、節口綱(せっこうこう)、腿口綱(たいこうこう)、およびメロストマ綱[3]Merostomata は、鋏角亜門に属する節足動物の分類群の1つ。総称として節口類、腿口類[3](merostome)がある。カブトガニ類とウミサソリ類を含むグループを形成している。
学名「Merostomata」はギリシャ語の「meros」(太もも)と「stoma」(口)の合成である。これは本群の動物において、歩脚の基部が口器のように特殊化したことに由来する[4]。
目次
1 用法
2 特徴
3 系統関係
4 該当する分類群
4.1 カブトガニ類
4.2 ウミサソリ類
4.3 Chasmataspidida類
4.4 その他のもの
5 脚注
6 参考文献
7 関連項目
8 外部リンク
用法
1852年、Merostomata はジェームズ・デーナによって創設され、当時はカブトガニ類のみを含む分類群であった。1866年、Henry Woodwardはこの分類群にウミサソリ類を追加して以降、節口類(Merostomata Woodward, 1866)はカブトガニ類とウミサソリ類を含んだ分類群として広く流用されている。
しかし、ウミサソリ類が後にクモガタ類の姉妹群とされることに至り、この分類群はクモガタ類を除いた側系統群とされるか、もしくはウミサソリ類を節口類から除外し、節口類(Merostomata Dana, 1852)をカブトガニ類 Xiphosura のシノニムとみなし[5]、この分類群は破棄すべき[6]、などの見解がある。なお、この分類群の単系統性を支持する研究結果もある[7]。
本項目では、カブトガニ類とウミサソリ類などを含む節口類について扱う。
特徴
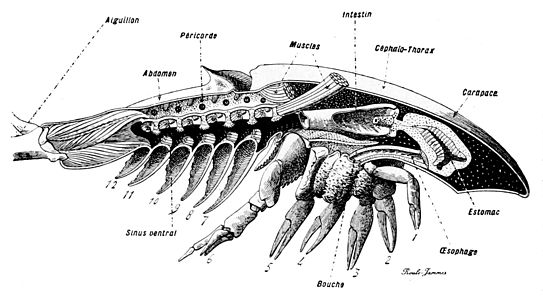
カブトガニ類の断面図


ウミサソリ類ユーリプテルスの体制模式図
節口類は、現生ではカブトガニ類のみを含む群であが、化石種は多く知られ、特に古生代の地層から多くの化石が見つかっている。大型種をも含む水生の節足動物である。
体は前体(prosoma)と後体(opisthosoma)、および尾節(telson)からなる[8]。
前体および頭胸部は、背面が広がって背甲(carapace)を構成する。通常、背甲の背面にはそれぞれ1対の複眼と単眼がある。6対の付属肢(関節肢)は全て背甲の下面に備える。最初の1対の付属肢は鋏角であり、通常は短く、先端は鋏となっている。次の5対は歩脚型の附属肢であり、最初のものは触肢であるが、歩脚から形態上の区別はほぼない[8]。両側の附属肢はその基部が正中線で接しており、基部の節は口器の役目をするように顎基(gnathobase)をもつものが多い。全ての鋏角類と同様、触角と真の顎はない。
後体(および腹部)は9-13節によって構成され[8]、現生種を含んだカブトガニ亜目では体節が互いに癒合しているが、それ以外の化石種では体節が区別される例が多い。腹面には附属肢があるが、鰭状や板状の蓋板(operculum)となっているものが多い。特に注目されるのは、往々にして退化的な「microtergite」("小さな体節")となる後体第1節(第7体節)である。この体節は外見上から完全に消失する例が多く、その体節の付属肢も多くの場合では退化的である[8]。ウェインベルギナはその例外で、第7体節の付属肢は前体のとほぼ一致な歩脚である[8]。
尾節は発達しており、棘状(尾剣)またはへら状となる。
一般に歩脚で海底を這い回る動物であるが、後体の鰭状付属肢や特殊化した遊泳脚で遊泳を行うものもある。
| 分類/体節 | 1 | 2-6 | 7 | 8 | 9-n |
|---|---|---|---|---|---|
カブトガニ亜目 | 鋏角 | 歩脚5対(後脚のみ二叉型) | 唇様肢 | 蓋板 | 蓋板(書鰓)5対 |
ウミサソリ類 † | 鋏角 | 歩脚5対 | Metastoma | 蓋板(生殖肢) | 蓋板(書鰓)4対 |
Chasmataspidida類 † | 鋏角? | 歩脚5対(二叉型?) | Metastoma | 蓋板(生殖肢) | 蓋板2対(5対?) |
ウェインベルギナ † | 鋏角 | 歩脚5対(二叉型?) | 歩脚 | 蓋板 | 蓋板5対? |
ディバステリウム † | 鋏角 | 歩脚5対(二叉型) | 唇様肢様の付属肢 | 蓋板(書鰓) | 蓋板(書鰓)5対 |
オッファコルス † | 鋏角 | 歩脚5対(二叉型) | 鰭状付属肢 | 蓋板 | 蓋板5対 |
クモガタ類 | 鋏角 | 触肢+歩脚4対 | -[9][8] | 蓋板(書肺)/- | 蓋板(書肺)/- |
ウミグモ類 | 鋏肢 | 触肢+担卵肢+歩脚3対 | 歩脚 | - | - |
系統関係

様々なクモガタ類
鋏角類の基本体制への考査において、節口類は特に注目されるグループである。これらの鋏角類からは、複眼・特殊化していない触肢・発達な後体付属肢・二叉型付属肢など、著しく特殊化したウミグモ類と派生的なクモガタ類には見当たらない、鋏角類の祖先形質(共有原始形質)と思われる特徴が少ながらずに発見できる。
従来、節口類はクモガタ類の姉妹群である単系統群と考えられた。しかしウミサソリ類については、のちに生殖器の構造に基づいてカブトガニ類よりもクモガタ類(クモ、サソリ、ダニなどが属する)に近縁の説が有力視される[5][8]。すなわち節口類は側系統群となり、ウミサソリ類とクモガタ類はSclerophorataという単系統群を構成する[5][8]。なお、少数であるものの、カブトガニ類とウミサソリ類からなる単系統群を支持する見解もある[7]。

かつては鋏角類と考えられた光楯類
光楯類はかつて鋏角類であると考えられ、カブトガニ類に分類された[5]。しかし再検討により、光楯類は鋏角類の前体付属肢構成を欠いている(鋏角はなく、歩脚も4-5対しかない)別系統の節足動物であると判明し、三葉虫類と共にArtiopoda類に分類されるようになった[10]。
下記の系統図は、Lamsdell et al. 2013[5]に基づいて簡略化したものである。クモガタ類以外の真鋏角類は、節口類に該当するグループである。また、Dekatriata以外のProsomapodaは、側系統群である広義のカブトガニ類に属する(後述参照)[5]。
| 鋏角亜門 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
該当する分類群
カブトガニ類

様々なカブトガニ類
カブトガニ類および剣尾類(カブトガニ目、剣尾目 Xiphosura)[11][12]は、現生は3属4種のみが生存し、化石はオルドビス紀まで遡り[13][1]、80種以上が認められるグループである[14]。ウミサソリ類に比べて体は比較的にコンパクトな形をとる。前体は歩脚を覆うドーム状の背甲であり、歩脚の先端が鋏になっているものが多く。後体は1対の生殖口蓋と5対の鰓脚をもつ。化石種は比較的に多様化したが、現生種は外見と生態とも大まかに共通している[15]。
狭義のカブトガニ類は、後体第1節(第7体節)は外見上から消失し、その付属肢は小さな唇様肢(chilaria)であることを共有形質とする[8]。広義のカブトガニ類は全てのSynziphosurida類をも含め、その中でも、第7体節の付属肢は歩脚であるという、他の真鋏角類に今まで発見例のない特徴をもったウェインベルギナがある[5]。カブトガニ亜目(Xiphosurida)の場合、後体の体節は全て癒合し、「thoracetron」という合体節となる[5][8]。名前の通り、長く伸びていた「尾剣」と呼ばれる剣状の尾節をもつ。
従来は単系統群とされ、カブトガニ亜目(Xiphosurida)とSynziphosurida亜目(Synziphosurine)の2群に細分された[16]。Lamsdell et al. 2013 の再検討により、Synziphosurida類は系統範囲の雑多な側系統群とされ、従来および広義のカブトガニ類は、ウミサソリ類やクモガタ類など(Dekatriata)を除いた側系統群とされる(カブトガニ類#系統関係および上記の系統図を参照)[5]。狭義のカブトガニ類は、カブトガニ亜目といくつかのSynziphosurida類のみを含んだ単系統群である[5]。
現生種は東アジアによるカブトガニ Tachypleus tridentatus 、南アジアと南東アジアによるミナミカブトガニ Tachypleus gigas とマルオカブトガニ Carcinoscorpius rotundicauda 、および北アメリカによるアメリカカブトガニ Limulus polyphemus の計4種が知られる。特にアメリカカブトガニの個体数は多くて研究も進んでいる[15]。鰓脚を用いて泳ぐことができ、雑食性で、二枚貝・巻貝・多毛類・甲殻類などの小動物や[17]、腐肉と海藻をも餌とする[18]。

Limuloides limuloides

ウェインベルギナ Weinbergina opitzi

ユウプループス Euproops rotundatus

オーストロリムルス Austrolimulus fletcheri

メソリムルス Mesolimulus walchi

アメリカカブトガニ Limulus polyphemus

カブトガニ Tachypleus tridentatus







Xiphosura sensu lato(広義のカブトガニ類、側系統群)[14]
Prosomapoda(Dekatriataを除く)- ウェインベルギナ科 Weinberginidae †
Planaterga(Dekatriataを除く)- (科)Bunodidae †
カブトガニ目 Xiphosura (狭義のカブトガニ類、単系統群)[5][14]- (科)Kasibelinuridae †
カブトガニ亜目 Xiphosurida- (下目)Bellinurina †
- (科)Bellinuridae †
- カブトガニ下目 Limulina
- (科)Rolfeiidae †
- (上科)Paleolimuloidea †
- (科)Paleolimulidae †
- カブトガニ上科 Limuloidea
オーストロリムルス科 Austrolimulidae †
カブトガニ科 Limulidae- (属)Limulus
アメリカカブトガニ Limulus polyphemus
- (亜科)Tachypleininae[15]
- (属)Tachypleus
カブトガニ Tachypleus tridentatus
ミナミカブトガニ Tachypleus gigas
- (属)Carcinoscorpius
マルオカブトガニ Carcinoscorpius rotundicauda
- (属)Tachypleus
- (属)Limulus
- (下目)Bellinurina †
- (科)Kasibelinuridae †
ウミサソリ類

様々なウミサソリ類
ウミサソリ類および広翼類(ウミサソリ目、広翼目 Eurypterida)は、およそ250種が認められ[14]、既知最大級の節足動物をも含んだ絶滅したグループである。カブトガニ類に比べて体は細長く、流線型の形をとる。前体は幅広いが、カブトガニ類ほどには極端でない。鋏角や歩脚が特殊化した例が存在し、特にユーリプテルス亜目はヘラ状の遊泳脚へ特殊化した後脚を持つ。後体は発達しており、13節のうち「metastoma」という癒合した小さな付属肢をもつ第1節は退化し、外見上は12節となる[8]。後体は更に前7節の中体と後5節の終体に区別される場合が多く、中体は特殊化した生殖肢と鰓のある蓋板によって構成される鰓室をもつ[8]。棘状ないしへら状の尾節をもつ[8]。
海中および淡水域に生息し、オルドビス紀からペルム紀まで繁栄した。化石から、海中からだんだんと淡水へと生息域を広めていったことがわかっている。
カブトガニ類の姉妹群とする見解が伝統的で、後にクモガタ類の姉妹群とする系統が有力視される[5][8][14]。

スティロヌルス Stylonurus powriensis

メガラシネ Megarachne servinei

ヒベルトプテルス Hibbertopterus scouleri

ミクソプテルス Mixopterus kiaeri

プテリゴトゥス Pterygotus anglicus

ユーリプテルス Eurypterus






ウミサソリ目 Eurypterida †
スティロヌルス亜目 Stylonurina †- (上科)Rhenopteroidea †(=Brachyopterellidae †)
- (科)Rhenopteridae †
スティロヌルス上科 Stylonuroidea †(=Drepanopteroidea †)
Stylonuridae †
Parastylonuridae †
ココモプテルス上科 Kokomopteroidea †
ココモプテルス科 Kokomopteridae †- (科)Hardieopteridae †
- (上科)Mycteropoidea †(=ヒベルトプテルス上科 Hibbertopteroidea †)
Drepanopteridae †
ヒベルトプテルス科 Hibbertopteridae †
Mycteroptidae †
- (上科)Rhenopteroidea †(=Brachyopterellidae †)
ユーリプテルス亜目 Eurypterina †- (上科)Moselopteroidea †
- (科)Moselopteridae †
カルシノソーマ上科 Carcinosomatoidea †(=メガログラプトゥス上科 Megalograptoidea †、ミクソプテルス上科 Mixopteroidea †)
カルシノソーマ科 Carcinosomatidae †
メガログラプトゥス科 Megalograptidae †
ミクソプテルス科 Mixopteridae †
ユーリプテルス上科 Eurypteroidea †- (科)Dolichopteridae †
- (科)Eurypteridae †
- (科)Strobilopteridae †
- (上科)Waeringopteroidea †(=Orcanopteridae †)
- (科)Waeringopteridae
- (科)Waeringopteridae
アデロフサルムス上科 Adelophthalmoidea †(=Nanahughmilleridae †、Parahughmilleriidae †)
アデロフサルムス科 Adelophthalmidae †
プテリゴトゥス上科 Pterygotioidea †(=スリモニア上科 Slimonioidea †、フグミレリア上科 Hughmillerioidea †)
フグミレリア科 Hughmilleriidae †
スリモニア科 Slimonidae †
プテリゴトゥス科 Pterygotidae †
- (上科)Moselopteroidea †
Chasmataspidida類
Chasmataspidida類(chasmataspids)は、11種が知られる[14]希少なグループである。いくつかの特徴はウミサソリに似通っているが、後体の体節分化は明らかに異なる[19][8]。鋏角の詳細は不明で、残りの前体付属肢も往々にして保存されていないが、知られるものからでは歩脚型で、Chasmataspis の場合は鋏型の先端をもつ[19]。Diploaspididae科の場合、後脚はへら状の遊泳脚となる[8]。後体は13節からなり、4節の中体と9節の終体という他の鋏角類に見当たらない独特な中-終体の組み合わせをもつ[19][8]。Chasmataspis の場合、中体の体節はカブトガニの後体のように癒合している[19]。中体はやや幅広く、第1節(第7体節)は消失しないもののごく短い[8]。少なくともDiploaspididaeの場合、腹側はウミサソリ類と同様に、「metastoma」という癒合した短い付属肢をもち、次の3節は蓋板をもち、そのうち最初のは生殖肢がある[8]。なお、この類は6対の蓋板をもったことを示唆するカンブリア紀のChasmataspis らしい生痕化石がある[19]。9節の終体は細長くなり、尾端は棘状ないしへら状の尾節をもつ[8]。
ウミサソリ類とクモガタ類からなるクレード(Sclerophorata)と姉妹群をなす系統[5]が有力視されるが、Chasmataspidida類の節口類における系統的位置は議論的である。特にこの類を側系統群とし、そのうちChasmataspis属(Chasmataspididae科)はカブトガニ類に近縁(カブトガニ類らしい背甲、鋏のある歩脚と部分的に癒合した後体に基づく)、Diploaspididae科はウミサソリ類に近縁(遊泳脚、生殖肢とmetastomaをもつなどの形質に基づく)であるという見解があった。しかし基盤的なカブトガニ類とウミサソリ類は、それぞれ分節した後体と歩脚型の後脚があり、前述の両者らしい形質はむしろ派生的で、収斂進化の結果であることを示唆する[19]。独特な中-終体の組み合わせも、Chasmataspidida類の単系統性を支持する共有派生形質と見なすことができる[19]。
- (目)Chasmataspidida †
- (科)Chasmataspididae †
- (属)Chasmataspis †
- (科)Diploaspididae †
- (属)Achanarraspis †
- (属)Diploaspis †
- (属)Dvulikiaspis †
- (属)Forfarella †
- (属)Heteroaspis †
- (属)Loganamaraspis †
- (属)Nahlyostaspis †
- (属)Octoberaspis †
- (属)Skrytyaspis †
- (科)Chasmataspididae †
その他のもの
以下の種類は、従来はカブトガニ類と考えられ、後に基盤的な真鋏角類と見なされるようになったものである[5]。カブトガニ類とウミサソリ類の場合、後脚以外の全ての歩脚は完全に外肢を欠いている単枝型であるが、これらの種類は、前体のほぼ全ての歩脚は二叉型で、よく発達した歩脚状の外肢を持っていた[8]。
オッファコルス Offacolus †
ディバステリウム Dibasterium †
脚注
- ^ abc“(PDF) A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids” (英語). ResearchGate. 2018年11月18日閲覧。
^ カンブリア紀まで遡る節口類らしい生痕化石がある。後述参照。- ^ ab小項目事典, ブリタニカ国際大百科事典. “腿口類(たいこうるい)とは - コトバンク” (日本語). コトバンク. 2018年11月11日閲覧。
^ “Virtual museum - Merostomata (Merostomates)”. muzeum.geology.cz. 2018年11月18日閲覧。- ^ abcdefghijklmnLamsdell, James C. (2012年12月18日). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura” (英語). Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x. ISSN 0024-4082. https://academic.oup.com/zoolinnean/article-abstract/167/1/1/2420794.
^ Tudge, Colin (2002) (英語). The Variety of Life: A Survey and a Celebration of All the Creatures that Have Ever Lived. Oxford University Press. ISBN 9780198604266. https://books.google.com/books?id=YW-2gnuU0L0C&pg=PA312.- ^ abGarwood, Russell J.; Dunlop, Jason (2014年11月13日). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359. https://peerj.com/articles/641/.
- ^ abcdefghijklmnopqrstuvA., Dunlop, Jason; C., Lamsdell, James. “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3). ISSN 1467-8039. https://www.academia.edu/28212892/Segmentation_and_tagmosis_in_Chelicerata.
^ サソリの前体の腹板(sternum)を第7体節の付属肢の名残とみなす説がある
^ Ortega-Hernández, Javier; Legg, David A.; Braddy, Simon J. (2012年8月15日). “The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda” (英語). Cladistics 29 (1): 15–45. doi:10.1111/j.1096-0031.2012.00413.x. ISSN 0748-3007. https://doi.org/10.1111/j.1096-0031.2012.00413.x.
^ 小項目事典, ブリタニカ国際大百科事典. “カブトガニ類(カブトガニるい)とは - コトバンク” (日本語). コトバンク. 2018年11月15日閲覧。
^ 第2版,日本大百科全書(ニッポニカ), ブリタニカ国際大百科事典 小項目事典,デジタル大辞泉,世界大百科事典. “剣尾類(けんびるい)とは - コトバンク” (日本語). コトバンク. 2018年11月15日閲覧。
^ The oldest horseshoe crab: A new xiphosurid from Late Ordovician Konservat-Lagerstatten deposits, Manitoba, Canada- ^ abcdefDunlop, J. A., Penney, D. & Jekel, D. 2015. A summary list of fossil spiders and their relatives.
- ^ abcDolejš, Petr; Vaňousová, Kateřina (2015年6月1日). A collection of horseshoe crabs (Chelicerata: Xiphosura) in the National Museum, Prague (Czech Republic) and a review of their immunological importance. 49. pp. 1–9. doi:10.5431/aramit4901. https://www.researchgate.net/publication/277711246_A_collection_of_horseshoe_crabs_Chelicerata_Xiphosura_in_the_National_Museum_Prague_Czech_Republic_and_a_review_of_their_immunological_importance.
^ ondrej.zicha(at)gmail.com, Ondrej Zicha;. “BioLib: Biological library” (チェコ語). www.biolib.cz. 2018年11月10日閲覧。
^ “(PDF) Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA” (英語). ResearchGate. 2018年11月15日閲覧。
^ “Horseshoe Crabs, Limulus polyphemus”. MarineBio.org. 2018年11月15日閲覧。- ^ abcdefgJason A. Dunlop, Lyall I. Anderson & Simon J. Braddy (2004年). “A redescription of Chasmataspis laurencii Caster & Brooks (Chelicerata: Chasmataspidida) from the Middle Ordovician of Tennessee, USA, with remarks on chasmataspid phylogeny” (PDF). Transactions of the Royal Society of Edinburgh: Earth Sciences 94 (4): 207–225. doi:10.1017/S0263593300000626. http://www.museunacional.ufrj.br/mndi/Aracnologia/Chasmataspidida/Dunlop%20et%20al%202007%20Chasmataspis.pdf.
参考文献
石川良輔編『節足動物の多様性と系統』,(2008),バイオディバーシティ・シリーズ6(裳華房)
関連項目
- 鋏角亜門
- カブトガニ類
- ウミサソリ
- Synziphosurine
- Chasmataspidida
- 生きている化石
外部リンク
- Merostomata - Tree | BioLib.cz
アメリカカブトガニ (英語)


